Biologia per il liceo/Invertebrati
Introduzione
modificaUna rapida occhiata a qualsiasi rivista relativa al nostro mondo naturale, come National Geographic , mostrerebbe una ricca varietà di vertebrati, in particolare mammiferi e uccelli. Per la maggior parte delle persone, questi sono gli animali che attraggono la nostra attenzione. Concentrarsi sui vertebrati, tuttavia, ci offre una visione piuttosto parziale e limitata della diversità animale, perché ignora quasi il 97 percento del regno animale, gli invertebrati, animali privi di cranio e di una colonna vertebrale o spina dorsale definita.
I phyla degli animali invertebrati presentano un'enorme varietà di cellule e tessuti adattati a scopi specifici, e spesso questi tessuti sono unici per i loro phyla. Queste specializzazioni mostrano la gamma di differenziazione cellulare possibile all'interno del clade Opisthokonta, che ha membri sia unicellulari che multicellulari. Le specializzazioni cellulari e strutturali includono cuticole per la protezione, spine e piccoli arpioni per la difesa, strutture dentate per l'alimentazione e ali per il volo. Un esoscheletro può essere adattato per il movimento o per l'attacco dei muscoli come nelle vongole e negli insetti. Le cellule secretorie possono produrre veleno, muco o enzimi digestivi. I piani corporei di alcuni phyla, come quelli dei molluschi, anellidi, artropodi ed echinodermi, sono stati modificati e adattati nel corso dell'evoluzione per produrre migliaia di forme diverse. Forse troverai sorprendente che un numero enorme di invertebrati sia acquatici che terrestri, forse milioni di specie, non siano ancora stati classificati scientificamente. Di conseguenza, le relazioni filogenetiche tra gli invertebrati vengono costantemente aggiornate man mano che vengono raccolte nuove informazioni sugli organismi di ciascun phylum.
Phylum dei Poriferi
modificaCome abbiamo visto, la stragrande maggioranza degli animali invertebrati non possiede un endoscheletro vertebrale osseo definito, o un cranio osseo. Tuttavia, uno dei gruppi più ancestrali di invertebrati deuterostomi, gli Echinodermi, produce minuscole "ossa" scheletriche chiamate ossicini che costituiscono un vero e proprio endoscheletro , o scheletro interno, ricoperto da un'epidermide.
Inizieremo la nostra indagine con il più semplice di tutti gli invertebrati: animali talvolta classificati all'interno del clade Parazoa ("accanto agli animali"). Questo clade attualmente include solo il phylum Placozoa (contenente una singola specie, Trichoplax adhaerens ), e il phylum Porifera , contenente le più familiari spugne ( Figura sotto). La divisione tra i Parazoa e gli Eumetazoa (tutti cladi animali sopra i Parazoa) è probabilmente avvenuta oltre un miliardo di anni fa.
Dovremmo ribadire qui che i Poriferi non possiedono tessuti "veri" che siano embriologicamente omologhi a quelli di tutti gli altri gruppi animali derivati come insetti e mammiferi. Questo perché non creano una vera gastrula durante l'embriogenesi e, di conseguenza, non producono un vero endoderma o ectoderma. Ma anche se non sono considerati dotati di veri tessuti, hanno cellule specializzate che svolgono funzioni specifiche come i tessuti (ad esempio, il "pinacoderma" esterno di una spugna agisce come la nostra epidermide). Quindi, funzionalmente , si può dire che i poriferi abbiano tessuti; tuttavia, questi tessuti probabilmente non sono embriologicamente omologhi ai nostri.
Le larve di spugna (ad esempio, parenchimula e anfiblastula) sono flagellate e in grado di nuotare; tuttavia, gli adulti sono immobili e trascorrono la loro vita attaccati a un substrato. Poiché l'acqua è vitale per le spugne per l'alimentazione, l'escrezione e lo scambio di gas, la loro struttura corporea facilita il movimento dell'acqua attraverso la spugna. Vari canali, camere e cavità consentono all'acqua di muoversi attraverso la spugna per consentire lo scambio di cibo e rifiuti nonché lo scambio di gas a quasi tutte le cellule del corpo.

Morfologia delle spugne
modificaEsistono almeno 5.000 specie di spugne nominate, probabilmente con migliaia di altre ancora da classificare. La morfologia delle spugne più semplici assume la forma di un cilindro irregolare con una grande cavità centrale, lo spongocele , che occupa l'interno del cilindro ( Figura sotto). L'acqua entra nello spongocele attraverso numerosi pori, o osti , che creano aperture nella parete del corpo. L'acqua che entra nello spongocele viene espulsa tramite una grande apertura comune chiamata osculum . Tuttavia, dovremmo notare che le spugne presentano una gamma di diversità nelle forme del corpo, tra cui variazioni nelle dimensioni e nella forma dello spongocele, nonché nel numero e nella disposizione delle camere di alimentazione all'interno della parete del corpo. In alcune spugne, più camere di alimentazione si aprono da uno spongocele centrale e in altre, diverse camere di alimentazione collegate tra loro possono trovarsi tra i pori di ingresso e lo spongocele.
Sebbene le spugne non presentino una vera organizzazione a strati di tessuto, hanno un certo numero di "tessuti" funzionali composti da diversi tipi di cellule specializzate per funzioni distinte. Ad esempio, le cellule epiteliali chiamate pinacociti formano il corpo più esterno, chiamato pinacodermi , che svolge una funzione protettiva simile a quella della nostra epidermide. Sparsi tra i pinacodermi ci sono gli osti che consentono l'ingresso dell'acqua nel corpo della spugna. Questi pori hanno dato alle spugne il loro nome di phylum Porifera, ovvero portatori di pori. In alcune spugne, gli osti sono formati da porociti , singole cellule a forma di tubo che agiscono come valvole per regolare il flusso di acqua nello spongocele. In altre spugne, gli osti sono formati da pieghe nella parete del corpo della spugna. Tra lo strato esterno e le camere di alimentazione della spugna c'è una sostanza gelatinosa chiamata mesoile , che contiene fibre collagene. Vari tipi di cellule risiedono all'interno del mesoile, tra cui gli amebociti , le "cellule staminali" delle spugne, e gli sclerociti , che producono materiali scheletrici. La consistenza gelatinosa del mesoile agisce come un endoscheletro e mantiene la morfologia tubulare delle spugne.
Le camere di alimentazione all'interno della spugna sono rivestite da coanociti ("cellule del collare"). La struttura di un coanocita è fondamentale per la sua funzione, che è quella di generare una corrente d'acqua diretta attraverso la spugna e di intrappolare e ingerire particelle di cibo microscopiche tramite fagocitosi. Queste cellule di alimentazione sono simili nell'aspetto ai coanoflagellati unicellulari (Protista). Questa somiglianza suggerisce che spugne e coanoflagellati sono strettamente correlati e probabilmente condividono un'ascendenza comune. Il corpo del coanocita è incorporato nel mesoile e contiene tutti gli organelli necessari per la normale funzione cellulare. Sporgendo nello "spazio aperto" all'interno della camera di alimentazione c'è un collare a maglie composto da microvilli con un singolo flagello al centro della colonna. Il battito dei flagelli di tutti i coanociti attira l'acqua nella spugna attraverso i numerosi osti, negli spazi rivestiti dai coanociti e infine fuori attraverso l'osculo (o gli osculi, se la spugna è costituita da una colonia di spugne attaccate). Le particelle di cibo, tra cui batteri trasportati dall'acqua e organismi unicellulari come alghe e vari protisti simili ad animali, vengono intrappolate dal collare a setaccio dei coanociti, scivolano verso il corpo della cellula e vengono ingerite tramite fagocitosi. I coanociti svolgono anche un'altra sorprendente funzione: possono differenziarsi in sperma per la riproduzione sessuale, momento in cui vengono staccati dal mesoile e lasciano la spugna con l'acqua espulsa attraverso l'osculo.
-
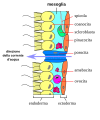 Rappresentazione schematica della parete di una spugna
Rappresentazione schematica della parete di una spugna -
 Schema di una spugna siconide
Schema di una spugna siconide -
 Struttura macroscopica dei Poriferi. A. tipo ascon B. tipo sycon C. tipo leucon pinacociti coanociti 1=spongocele; 2=osculo; 3=tubo radiale; 4=camera coanocitaria; 5=pori; 6=canali inalanti.
Struttura macroscopica dei Poriferi. A. tipo ascon B. tipo sycon C. tipo leucon pinacociti coanociti 1=spongocele; 2=osculo; 3=tubo radiale; 4=camera coanocitaria; 5=pori; 6=canali inalanti.



Video
modificaGuarda questo video per vedere il movimento dell'acqua attraverso il corpo della spugna.
Gli amebociti (derivati da archeociti simili a cellule staminali), sono così chiamati perché si muovono attraverso il mesoilo in modo simile alle amebe. Hanno una varietà di funzioni: oltre a fornire nutrienti dai coanociti ad altre cellule all'interno della spugna, danno origine anche alle uova per la riproduzione sessuale. (Le uova rimangono nel mesoilo, mentre gli spermatozoi vengono rilasciati nell'acqua.) Gli amebociti possono differenziarsi in altri tipi di cellule della spugna, come i collenciti e i lofociti, che producono la proteina simile al collagene che supporta il mesoilo. Gli amebociti possono anche dare origine agli sclerociti, che producono spicole (spighe scheletriche di silice o carbonato di calcio) in alcune spugne, e agli spongociti, che producono la proteina spongina nella maggior parte delle spugne. Questi diversi tipi di cellule nelle spugne sono mostrati nella Figura sotto.

Domanda: quale delle seguenti affermazioni è falsa?
- I coanociti sono dotati di flagelli che spingono l'acqua attraverso il corpo.
- I pinacociti possono trasformarsi in qualsiasi tipo di cellula.
- I lofociti secernono collagene.
- I porociti controllano il flusso dell'acqua attraverso i pori del corpo della spugna.
Video
modificaOsserva da vicino la spugna e le sue cellule.
Come abbiamo visto, la maggior parte delle spugne è sostenuta da piccole spicole ossee (solitamente piccole strutture appuntite fatte di carbonato di calcio o silice) nel mesoile. Le spicole forniscono supporto al corpo della spugna e possono anche scoraggiare la predazione. La presenza e la composizione delle spicole costituiscono la base per differenziare tre delle quattro classi di spugne ( Figura sotto). Le spugne della classe Calcarea producono spicole di carbonato di calcio e nessuna spongina; quelle della classe Hexactinellida producono spicole silicee (vetrose) a sei raggi e nessuna spongina; e quelle della classe Demospongia contengono spongina e possono o meno avere spicole; se presenti, quelle spicole sono silicee. Le spugne di quest'ultima classe sono state utilizzate come spugne da bagno. Le spicole sono presenti in modo più evidente nelle spugne di vetro, classe Hexactinellida. Alcune delle spicole possono raggiungere proporzioni gigantesche. Ad esempio, rispetto alle tipiche spicole di spugna di vetro, le cui dimensioni variano generalmente da 3 a 10 mm, alcune delle spicole basali dell'esattino Monorhaphis chuni sono enormi e crescono fino a 3 metri di lunghezza! Le spugne di vetro sono anche insolite in quanto la maggior parte delle loro cellule corporee sono fuse insieme per formare un sincizio multinucleato . Poiché le loro cellule sono interconnesse in questo modo, le spugne esattino non hanno mesoile. Una quarta classe di spugne, le Sclerospongiae, è stata descritta da specie scoperte in tunnel sottomarini. Queste sono anche chiamate spugne coralline per i loro scheletri multistrato di carbonato di calcio. La datazione basata sulla velocità di deposizione degli strati scheletrici suggerisce che alcune di queste spugne hanno centinaia di anni.
| a | b | c |
|---|---|---|
 |
 |
 |
| Diverse classi di spugne. (a) Clathrina clathrus appartiene alla classe Calcarea, (b) Staurocalyptus spp. (nome comune: spugna gialla Picasso) appartiene alla classe Hexactinellida, e (c) Acarnus erithacus appartiene alla classe Demospongia. | ||
Collegamento all'apprendimento
modificaUtilizza la Guida interattiva alle spugne per identificare le specie di spugne in base alla loro forma esterna, allo scheletro minerale, alle fibre e all'architettura scheletrica.
Processi fisiologici nelle spugne
modificaLe spugne, pur essendo organismi semplici, regolano i loro diversi processi fisiologici attraverso una varietà di meccanismi. Questi processi regolano il loro metabolismo, la riproduzione e la locomozione.
Digestione
modificaLe spugne non hanno sistemi digestivi, respiratori, circolatori e nervosi complessi. Il loro cibo viene intrappolato mentre l'acqua passa attraverso gli osti e fuoriesce attraverso l'osculo. I batteri di dimensioni inferiori a 0,5 micron vengono intrappolati dai coanociti, che sono le principali cellule impegnate nell'alimentazione, e vengono ingeriti tramite fagocitosi. Tuttavia, le particelle più grandi degli osti possono essere fagocitate sulla superficie della spugna dai pinacociti. In alcune spugne, gli amebociti trasportano il cibo dalle cellule che hanno ingerito particelle di cibo a quelle che non lo hanno fatto. Nelle spugne, nonostante quella che sembra una grande cavità digestiva, tutta la digestione è intracellulare . Il limite di questo tipo di digestione è che le particelle di cibo devono essere più piccole delle singole cellule della spugna.
Tutte le altre principali funzioni corporee nella spugna (scambio di gas, circolazione, escrezione) vengono eseguite per diffusione tra le cellule che rivestono le aperture all'interno della spugna e l'acqua che passa attraverso tali aperture. Tutti i tipi di cellule all'interno della spugna ottengono ossigeno dall'acqua tramite diffusione. Allo stesso modo, l'anidride carbonica viene rilasciata nell'acqua di mare tramite diffusione. Inoltre, i rifiuti azotati prodotti come sottoprodotto del metabolismo proteico vengono escreti tramite diffusione dalle singole cellule nell'acqua mentre passa attraverso la spugna.
Alcune spugne ospitano alghe verdi o cianobatteri come endosimbionti all'interno di archeociti e altre cellule. Potrebbe sorprendere scoprire che ci sono quasi 150 specie di spugne carnivore, che si nutrono principalmente di piccoli crostacei, intrappolandoli attraverso fili appiccicosi o spicole uncinate!
Sebbene le spugne non abbiano un sistema nervoso specializzato, esiste una comunicazione intercellulare in grado di regolare eventi come la contrazione del corpo della spugna o l'attività dei coanociti.
Riproduzione
modificaLe spugne si riproducono sia per via sessuale che asessuata. Il mezzo tipico della riproduzione asessuata è la frammentazione (durante questo processo, un pezzo di spugna si stacca, si deposita su un nuovo substrato e si sviluppa in un nuovo individuo) o la gemmazione (una crescita geneticamente identica cresce dal genitore e alla fine si stacca o rimane attaccata per formare una colonia). Un tipo atipico di riproduzione asessuata si trova solo nelle spugne d'acqua dolce e avviene attraverso la formazione di gemmule . Le gemmule sono strutture resistenti all'ambiente prodotte da spugne adulte (ad esempio, nella spugna d'acqua dolce Spongilla ). Nelle gemmule, uno strato interno di archeociti (amebociti) è circondato da uno strato cellulare pneumatico che può essere rinforzato con spicole. Nelle spugne d'acqua dolce, le gemmule possono sopravvivere a condizioni ambientali ostili come i cambiamenti di temperatura e quindi servire per ricolonizzare l'habitat una volta che le condizioni ambientali migliorano e si stabilizzano. Le gemmule sono in grado di attaccarsi a un substrato e generare una nuova spugna. Poiché le gemmule possono resistere ad ambienti difficili, sono resistenti alla disidratazione e rimangono dormienti per lunghi periodi, rappresentano un ottimo mezzo di colonizzazione per un organismo sessile.
La riproduzione sessuale nelle spugne avviene quando vengono generati i gameti. Gli ovociti nascono dalla differenziazione degli amebociti e vengono trattenuti all'interno dello spongocele, mentre gli spermatozoi derivano dalla differenziazione dei coanociti e vengono espulsi tramite l'osculum. Le spugne sono monoiche (ermafrodite), il che significa che un individuo può produrre entrambi i gameti (uova e sperma) contemporaneamente. In alcune spugne, la produzione di gameti può avvenire durante tutto l'anno, mentre altre spugne possono mostrare cicli sessuali a seconda della temperatura dell'acqua. Le spugne possono anche diventare ermafrodite in sequenza , producendo prima gli ovociti e poi gli spermatozoi. Questa separazione temporale dei gameti prodotti dalla stessa spugna aiuta a incoraggiare la fecondazione incrociata e la diversità genetica. Gli spermatozoi trasportati dalle correnti d'acqua possono fecondare gli ovociti trasportati nel mesoile di altre spugne. Lo sviluppo larvale precoce avviene all'interno della spugna e le larve che nuotano liberamente (come i parenchimi flagellati ) vengono poi rilasciate attraverso l'osculo.
Locomozione
modificaLe spugne sono generalmente sessili da adulte e trascorrono la loro vita attaccate a un substrato fisso. Non mostrano movimento su grandi distanze come altri invertebrati marini che nuotano liberamente. Tuttavia, le cellule delle spugne sono in grado di strisciare lungo i substrati tramite plasticità organizzativa , ovvero riorganizzando le loro cellule. In condizioni sperimentali, i ricercatori hanno dimostrato che le cellule delle spugne sparse su un supporto fisico dimostrano un vantaggio per il movimento diretto. Si è ipotizzato che questo movimento di strisciamento localizzato possa aiutare le spugne ad adattarsi ai microambienti vicino al punto di attacco. Va notato, tuttavia, che questo modello di movimento è stato documentato in laboratorio, ma deve ancora essere osservato negli habitat naturali delle spugne.
Collegamento all'apprendimento
modificaGuarda questo video della BBC che mostra la varietà di spugne osservate lungo il Cayman Wall durante un'immersione in immersione.
Phylum Cnidaria
modificaIl phylum Cnidaria comprende animali che presentano simmetria radiale o biradiale e sono diploblastici, ovvero si sviluppano da due strati embrionali, ectoderma ed endoderma. Quasi tutti (circa il 99%) gli cnidari sono specie marine.
Mentre il tipo di cellula che definisce le spugne è il coanocita, il tipo di cellula che definisce gli cnidari è lo cnidocita , o cellula urticante. Queste cellule si trovano attorno alla bocca e sui tentacoli e servono a catturare le prede o a respingere i predatori. Gli cnidociti hanno grandi organelli urticanti chiamati nematocisti , che di solito contengono punte alla base di un lungo filo arrotolato. La parete esterna della cellula ha una proiezione simile a un capello chiamata cnidocillo , che è sensibile alla stimolazione tattile. Se i cnidocilli vengono toccati, i fili cavi si rovesciano con un'enorme accelerazione, che si avvicina a 40.000 volte quella della gravità. I fili microscopici quindi aggrovigliano la preda o penetrano istantaneamente nella carne della preda o del predatore, rilasciando tossine (tra cui neurotossine e tossine che formano pori che possono portare alla lisi cellulare) nel bersaglio, immobilizzandolo o paralizzandolo (vedere Figura sotto).

Video
modificaGuarda questa animazione video che mostra due anemoni impegnati in una battaglia.
Nei Cnidari si trovano due distinti piani corporei: la forma a polipo o a tulipano "a gambo" e la forma a medusa o "a campana". ( Figura 28.6 ). Un esempio della forma a polipo si trova nel genere Hydra , mentre la forma più tipica di medusa si trova nel gruppo chiamato "meduse di mare" (meduse). Le forme a polipo sono sessili da adulte, con una singola apertura (la bocca/ano) verso la cavità digestiva rivolta verso l'alto con tentacoli che la circondano. Le forme a medusa sono mobili, con la bocca e i tentacoli che pendono da una campana a forma di ombrello.

Alcuni cnidari sono dimorfici , ovvero presentano entrambi i piani corporei durante il loro ciclo vitale. In queste specie, il polipo funge da fase asessuata, mentre la medusa funge da fase sessuale e produce gameti. Tuttavia, entrambe le forme corporee sono diploidi.
Un esempio di dimorfismo cnidarico può essere osservato nell'idroide coloniale Obelia . La colonia asessuata sessile ha due tipi di polipi, mostrati nella Figura sotto. Il primo è il gastrozoide , che è adattato per catturare prede e nutrirsi. In Obelia , tutti i polipi sono collegati tramite una cavità digestiva comune chiamata cenosarco . L'altro tipo di polipo è il gonozoide , adattato per la gemmazione asessuata e la produzione di meduse sessuali. Le gemme riproduttive del gonozoide si staccano e maturano in meduse che nuotano liberamente, che sono maschili o femminili (dioiche). Ogni medusa ha diversi testicoli o diverse ovaie in cui avviene la meiosi per produrre spermatozoi o cellule uovo. È interessante notare che le cellule che producono gameti non nascono all'interno della gonade stessa, ma migrano in essa dai tessuti del gonozoide. Questa origine separata di gonadi e gameti è comune in tutti gli eumetazoi. I gameti vengono rilasciati nell'acqua circostante e, dopo la fecondazione, lo zigote si sviluppa in una blastula, che presto si sviluppa in una larva planula ciliata e bilateralmente simmetrica. La planula nuota liberamente per un po', ma alla fine si attacca a un substrato e diventa un singolo polipo, da cui si forma una nuova colonia di polipi per gemmazione.

Video
modificaClicca qui per seguire l' animazione del ciclo di vita di Obelia .
Tutti gli cnidari sono diploblastici e quindi hanno due strati "epiteliali" nel corpo che derivano dall'endoderma e dall'ectoderma dell'embrione. Lo strato esterno (dall'ectoderma) è chiamato epidermide e riveste l'esterno dell'animale, mentre lo strato interno (dall'endoderma) è chiamato gastroderma e riveste la cavità digestiva. Nella larva planula, uno strato di ectoderma circonda una massa solida di endoderma, ma man mano che il polipo si sviluppa, la cavità digestiva o gastrovascolare si apre all'interno dell'endoderma. Una mesoglea gelatinosa non vivente si trova tra questi due strati epiteliali. In termini di complessità cellulare, gli cnidari mostrano la presenza di tipi di cellule differenziate in ogni strato di tessuto, come cellule nervose, cellule epiteliali contrattili, cellule che secernono enzimi e cellule che assorbono nutrienti, nonché la presenza di connessioni intercellulari. Tuttavia, a parte alcune notevoli eccezioni come statocisti e ropali (vedi sotto), lo sviluppo di organi o sistemi di organi non è avanzato in questo phylum.
Il sistema nervoso è rudimentale, con cellule nervose organizzate in una rete sparsa in tutto il corpo. Questa rete nervosa può mostrare la presenza di gruppi di cellule che formano plessi nervosi (singolare: plesso) o cordoni nervosi. L'organizzazione del sistema nervoso nella medusa mobile è più complessa di quella del polipo sessile, con un anello nervoso attorno al bordo della campana della medusa che controlla l'azione dei tentacoli. Le cellule nervose degli cnidari mostrano caratteristiche miste di neuroni motori e sensoriali. Le molecole di segnalazione predominanti in questi sistemi nervosi primitivi sono i peptidi, che svolgono sia funzioni eccitatorie che inibitorie. Nonostante la semplicità del sistema nervoso, è notevole che coordini il complicato movimento dei tentacoli, l'attrazione della preda catturata verso la bocca, la digestione del cibo e l'espulsione dei rifiuti.
La cavità gastrovascolare ha una sola apertura che funge sia da bocca che da ano; questa disposizione è chiamata sistema digerente incompleto. Nella cavità gastrovascolare, la digestione extracellulare avviene quando il cibo viene introdotto nella cavità gastrovascolare, gli enzimi vengono secreti nella cavità e le cellule che rivestono la cavità assorbono i nutrienti. Tuttavia, si verifica anche una certa digestione intracellulare. La cavità gastrovascolare distribuisce i nutrienti in tutto il corpo dell'animale, con i nutrienti che passano dalla cavità digestiva attraverso la mesoglea alle cellule epidermiche. Pertanto, questa cavità svolge sia funzioni digestive che circolatorie.
Le cellule degli cnidari scambiano ossigeno e anidride carbonica per diffusione tra le cellule nell'epidermide e l'acqua nell'ambiente, e tra le cellule nel gastroderma e l'acqua nella cavità gastrovascolare. La mancanza di un sistema circolatorio per spostare i gas disciolti limita lo spessore della parete corporea e necessita di una mesoglea non vivente tra gli strati. Negli cnidari con una mesoglea più spessa, un certo numero di canali aiuta a distribuire sia i nutrienti che i gas. Non c'è né un sistema escretore né organi, e i rifiuti azotati semplicemente si diffondono dalle cellule nell'acqua esterna all'animale o nella cavità gastrovascolare.
Il phylum Cnidaria contiene circa 10.000 specie descritte divise in due cladi monofiletici: gli Anthozoa e i Medusozoa. Gli Anthozoa includono i coralli, le gorgonie, le fruste di mare e gli anemoni di mare. I Medusozoa includono diverse classi di Cnidaria in due cladi: gli Hydrozoa includono forme sessili, alcune forme medusoidi e forme coloniali natanti come la caravella portoghese. L'altro clade contiene vari tipi di meduse tra cui sia gli Scyphozoa che i Cubozoa. Gli Anthozoa contengono solo forme di polipo sessili, mentre i Medusozoa includono specie con forme sia di polipo che di medusa nel loro ciclo vitale.
Classe Antozoi
modificaLa classe Anthozoa ("animali dei fiori") comprende anemoni di mare ( Figura sotto ), penne di mare e coralli, con un numero stimato di 6.100 specie descritte. Gli anemoni di mare sono solitamente dai colori vivaci e possono raggiungere una dimensione di 1,8-10 cm di diametro. I singoli animali hanno una forma cilindrica e sono attaccati direttamente a un substrato.
| a | b |
|---|---|
 |
 |
| Anemone di mare. L'anemone di mare è mostrato (a) fotografato e (b) in un diagramma che ne illustra la morfologia. | |
La bocca di un anemone di mare è circondata da tentacoli che portano cnidociti. L'apertura della bocca a fessura e la faringe appiattita sono rivestite di ectoderma. Questa struttura della faringe rende gli anemoni bilateralmente simmetrici. Un solco ciliato chiamato sifonoglifo si trova su due lati opposti della faringe e dirige l'acqua al suo interno. La faringe è la parte muscolare dell'apparato digerente che serve per ingerire e egerire il cibo e può estendersi fino a due terzi della lunghezza del corpo prima di aprirsi nella cavità gastrovascolare. Questa cavità è divisa in diverse camere da setti longitudinali chiamati mesenteri . Ogni mesentere è costituito da una piega di tessuto gastrodermico con uno strato di mesoglea tra gli strati di gastroderma. I mesenteri non dividono completamente la cavità gastrovascolare e le cavità più piccole si fondono all'apertura faringea. Il vantaggio adattivo dei mesenteri sembra essere un aumento della superficie per l'assorbimento dei nutrienti e lo scambio di gas, nonché un ulteriore supporto meccanico per il corpo dell'anemone.
Gli anemoni di mare si nutrono di piccoli pesci e gamberetti, solitamente immobilizzando le loro prede con nematocisti. Alcuni anemoni di mare stabiliscono una relazione mutualistica con i paguri quando il granchio li cattura e li attacca alla loro conchiglia. In questa relazione, l'anemone ottiene particelle di cibo dalle prede catturate dal granchio, e il granchio è protetto dai predatori dalle cellule urticanti dell'anemone. Alcune specie di pesce pagliaccio, o pesce pagliaccio, sono anche in grado di vivere con gli anemoni di mare perché sviluppano un'immunità acquisita alle tossine contenute nei nematocisti e secernono anche un muco protettivo che impedisce loro di essere punti.
La struttura dei polipi dei coralli è simile a quella degli anemoni, sebbene i singoli polipi siano solitamente più piccoli e facciano parte di una colonia, alcuni dei quali sono enormi e delle dimensioni di piccoli edifici. I polipi dei coralli si nutrono di organismi planctonici più piccoli, tra cui alghe, batteri e larve di invertebrati. Alcuni antozoi hanno associazioni simbiotiche con alghe dinoflagellate chiamate zooxantelle. La relazione reciprocamente vantaggiosa tra zooxantelle e coralli moderni, che fornisce riparo alle alghe, conferisce alle barriere coralline i loro colori e fornisce nutrienti a entrambi gli organismi. Questa complessa associazione mutualistica è iniziata più di 210 milioni di anni fa, secondo un nuovo studio di un team internazionale di scienziati. Il fatto che questa relazione simbiotica sia nata durante un periodo di massiccia espansione delle barriere coralline in tutto il mondo suggerisce che l'interconnessione tra alghe e coralli è fondamentale per la salute delle barriere coralline, che forniscono habitat a circa un quarto di tutta la vita marina. Le barriere coralline sono minacciate dal riscaldamento degli oceani, che ha portato i coralli a espellere le loro alghe zooxantelle e a diventare bianchi, un processo chiamato sbiancamento dei coralli.
Gli antozoi rimangono polipoidi (si noti che questo termine è facilmente confuso con "poliploide") per tutta la vita e possono riprodursi asessualmente per gemmazione o frammentazione, o sessualmente producendo gameti. I gameti maschili o femminili prodotti da un polipo si fondono per dare origine a una larva planula che nuota liberamente. La larva si insedia su un substrato adatto e si sviluppa in un polipo sessile.
Classe Scyphozoa
modificaLa classe Scyphozoa ("animali a coppa") include solo meduse marine, con circa 200 specie note. La medusa è lo stadio principale nel ciclo vitale, sebbene vi sia uno stadio di polipo nel ciclo vitale della maggior parte delle specie. La maggior parte delle meduse varia da 2 a 40 cm di lunghezza, ma la specie di scyphozoa più grande, Cyanea capillata , può raggiungere una dimensione di due metri di diametro. Gli scyphozoi mostrano una caratteristica morfologia a campana ( Figura sotto).
| a | b |
|---|---|
 |
 |
| Una medusa marina. Una medusa è mostrata (a) fotografata e (b) in un diagramma che ne illustra la morfologia | |

Nella medusa marina, è presente un'apertura boccale sulla parte inferiore dell'animale, circondata da tentacoli cavi che portano nematocisti. Gli scifozoi vivono la maggior parte del loro ciclo vitale come carnivori solitari e nuotatori liberi. La bocca conduce alla cavità gastrovascolare, che può essere sezionata in quattro sacche interconnesse, chiamate diverticoli . In alcune specie, l'apparato digerente può ramificarsi ulteriormente in canali radiali . Come i setti negli antozoi, le cellule gastrovascolari ramificate svolgono due funzioni: aumentare la superficie per l'assorbimento e la diffusione dei nutrienti e sostenere il corpo dell'animale.
Negli scifozoi, le cellule nervose sono organizzate in una rete nervosa che si estende su tutto il corpo, con un anello nervoso attorno al bordo della campana. Gruppi di organi sensoriali chiamati rhopalia possono essere presenti in tasche sul bordo della campana. Le meduse hanno un anello di muscoli che riveste la cupola del corpo, che fornisce la forza contrattile necessaria per nuotare nell'acqua, nonché per estrarre il cibo dall'acqua mentre nuotano. Gli scifozoi hanno sessi separati. Le gonadi sono formate dal gastroderma e i gameti vengono espulsi attraverso la bocca. Le larve di planula sono formate da fecondazione esterna; si stabiliscono su un substrato in una forma polipoide. Questi polipi possono gemmare per formare altri polipi o iniziare immediatamente a produrre gemme di medusa. In alcune specie, la larva di planula può svilupparsi direttamente nella medusa. Il ciclo di vita ( Figura sotto) della maggior parte degli scifozoi include sia forme corporee medusoidi sessuate che polipoidi asessuate.

Classe Cubozoi
modificaQuesta classe include meduse che hanno una medusa a forma di scatola, o una campana di sezione trasversale quadrata, e sono colloquialmente note come "meduse scatola". Queste specie possono raggiungere dimensioni da 15 a 25 cm, ma in genere i membri dei Cubozoa non sono grandi quanto quelli degli Scyphozoa. Tuttavia, i cubozoi mostrano caratteristiche morfologiche e anatomiche complessive simili a quelle degli Scyphozoi. Una differenza importante tra le due classi è la disposizione dei tentacoli. I cubozoi contengono cuscinetti muscolari chiamati pedalia agli angoli della calotta a campana quadrata, con uno o più tentacoli attaccati a ciascun pedalio. In alcuni casi, l'apparato digerente può estendersi nella pedalia. I nematocisti possono essere disposti in una configurazione a spirale lungo i tentacoli; questa disposizione aiuta a sottomettere e catturare efficacemente la preda. I cubozoi includono i più velenosi di tutti gli cnidari ( Figura sotto).
Questi animali sono insoliti perché hanno occhi che formano immagini, tra cui una cornea, un cristallino e una retina. Poiché queste strutture sono fatte da una serie di tessuti interattivi, possono essere chiamate veri e propri organi . Gli occhi sono situati in quattro gruppi tra ogni coppia di pedali. Ogni gruppo è costituito da quattro semplici macchie oculari più due occhi che formano immagini orientati in direzioni diverse. Il modo in cui le immagini formate da questi occhi molto complessi vengono elaborate rimane un mistero, poiché i cubozoi hanno ampie reti nervose ma nessun cervello distinto. Tuttavia, la presenza di occhi aiuta i cubozoi a essere cacciatori attivi ed efficaci di piccoli animali marini come vermi, artropodi e pesci.
I cubozoi hanno sessi separati e la fecondazione avviene all'interno della femmina. Le larve di planula possono svilupparsi all'interno della femmina o essere rilasciate, a seconda della specie. Ogni planula si sviluppa in un polipo. Questi polipi possono germogliare per formare altri polipi e creare una colonia; ogni polipo si trasforma quindi in una singola medusa.
| a | b | c |
|---|---|---|
 |
 |
 |
| n cubozoo. La (a) minuscola gelatina cubozoica Malo kingi ha la forma di un ditale e, come tutte le gelatine cubozoiche, (b) ha quattro pedali muscolari a cui si attaccano i tentacoli. La M. kingi è una delle due specie di gelatine note per causare la sindrome di Irukandji, una condizione caratterizzata da lancinanti dolori muscolari, vomito, aumento della frequenza cardiaca e sintomi psicologici. Si ritiene che due persone in Australia, dove le meduse Irukandji si trovano più comunemente, siano morte per punture di Irukandji. (c) Un cartello su una spiaggia australiana avverte i bagnanti del pericolo. | ||
Classe Idrozoi
modificaHydrozoa è un gruppo eterogeneo che comprende circa 3.200 specie; la maggior parte sono marine, anche se sono note alcune specie di acqua dolce ( Figura 28.12 ). La maggior parte delle specie presenta sia forme polipoidi che medusoidi nei loro cicli di vita, anche se la familiare Hydra ha solo la forma a polipo. La forma medusoide ha un velo muscolare o velo sotto il margine della campana e per questo motivo è chiamata idromedusa . Al contrario, la forma medusoide di Scyphozoa è priva di velo ed è chiamata scifomedusa .
La forma del polipo in questi animali mostra spesso una morfologia cilindrica con una cavità gastrovascolare centrale rivestita dal gastroderma. Il gastroderma e l'epidermide hanno uno strato semplice di mesoglea incastrato tra loro. Un'apertura della bocca, circondata da tentacoli, è presente all'estremità orale dell'animale. Molti idrozoi formano colonie sessili e ramificate di polipi specializzati che condividono una cavità gastrovascolare comune e ramificata (cenosarco), come si trova nell'idroide coloniale Obelia .
Le specie coloniali galleggianti chiamate sifonofori contengono individui sia medusoidi che polipoidi specializzati per l'alimentazione, la difesa o la riproduzione. Il caratteristico galleggiante color arcobaleno della caravella portoghese ( Physalia physalis ) crea uno pneumatoforo con cui regola la galleggiabilità riempiendola ed espellendo gas monossido di carbonio. A prima vista, questi complessi superorganismi sembrano essere un singolo organismo; ma la realtà è che persino i tentacoli sono in realtà composti da zooidi carichi di nematocisti. Quindi, sebbene superficialmente assomigli a una tipica medusa medusozoa, la P. physalis è una colonia di idrozoi galleggianti ; ogni esemplare è composto da molte centinaia di organismi, ciascuno specializzato per una determinata funzione, tra cui motilità e galleggiabilità, alimentazione, riproduzione e difesa. Sebbene siano carnivori e si nutrano di molti animali marini dal corpo molle, i P. physalis non hanno stomaco e hanno invece polipi specializzati chiamati gastrozoidi che usano per digerire le prede in acque libere.
Physalia ha colonie maschili e femminili, che rilasciano i loro gameti nell'acqua. Lo zigote si sviluppa in un singolo individuo, che poi germoglia asessualmente per formare una nuova colonia. I sifonofori includono le più grandi colonie di cnidari galleggianti conosciute come Praya dubia , la cui catena di zooidi può arrivare fino a 50 metri (165 piedi) di lunghezza. Altre specie di idrozoi sono polipi solitari ( Hydra ) o idromeduse solitarie ( Gonionemus ). Una caratteristica distintiva condivisa dagli idrozoi è che le loro gonadi derivano dal tessuto epidermico, mentre in tutti gli altri cnidari derivano dal tessuto gastrodermico.
| a | b |
|---|---|
 |
 |
| c | d |
 |
 |
| Idrozoi. La Tubularia indivisa (a), le colonie di sifonofori Physalia physalis , nota come caravella portoghese (b), Velella bae (c) e il polipo solitario Hydra (d) hanno forme corporee diverse, ma appartengono tutte alla famiglia degli Idrozoi. | |
Superphylum Lophotrochozoa: vermi piatti, rotiferi e nemertei
modificaGli animali appartenenti al superphylum Lophotrochozoa sono triploblastici (hanno tre foglietti germinali) e, a differenza degli cnidari, possiedono un mesoderma embrionale inserito tra l'ectoderma e l'endoderma. Questi phyla sono anche bilateralmente simmetrici, il che significa che una sezione longitudinale li dividerà in lati destro e sinistro che sono superficialmente simmetrici. In questi phyla, vediamo anche l'inizio della cefalizzazione, l'evoluzione di una concentrazione di tessuti nervosi e organi sensoriali nella testa dell'organismo, esattamente dove un organismo mobile bilateralmente simmetrico incontra per la prima volta il suo ambiente.
I lofotrocozoi sono anche protostomi, in cui il blastoporo, o il punto di invaginazione dell'ectoderma (strato germinale esterno), diventa l'apertura della bocca nel canale alimentare. Questo modello di sviluppo è chiamato protostomia o "prima bocca". I protostomi includono i phyla acelomato, pseudocelomato ed eucelomato. Il celoma è una cavità che separa l'ectoderma dall'endoderma. Negli acelomati, una massa solida di mesoderma è inserita tra l'ectoderma e l'endoderma e non forma una cavità o "celoma", lasciando poco spazio per lo sviluppo degli organi; negli pseudocelomati, c'è una cavità o pseudoceloma che sostituisce il blastocele (la cavità all'interno della blastula), ma è rivestita dal mesoderma solo all'esterno della cavità, lasciando il tubo intestinale e gli organi non rivestiti; negli eucelomati, la cavità che oblitera il blastocele man mano che si sviluppa il celoma è rivestita sia all'esterno della cavità (strato parietale) sia all'interno della cavità, attorno al tubo intestinale e agli organi interni (strato viscerale).
I protostomi eucoelmati sono schizocele, in cui le cellule che producono mesoderma migrano tipicamente nel blastocele durante la gastrulazione e si moltiplicano per formare una massa solida di cellule. Quindi si sviluppano delle cavità all'interno della massa cellulare per formare il celoma. Poiché la cavità corporea in formazione divide il mesoderma, questo celoma protostomico è definito schizocele . Come vedremo più avanti in questo capitolo, i cordati, il phylum a cui apparteniamo, generalmente sviluppano un celoma tramite enterocele: sacche di mesoderma pizzicano l'intestino primitivo invaginante, o archenteron , e poi si fondono per formare un celoma completo. Dovremmo notare qui che un eucoelomato può formare il suo "vero celoma" tramite schizocele o enterocele. Il processo che produce il celoma è diverso e di importanza tassonomica, ma il risultato è lo stesso: un celoma completo, rivestito di mesoderma.
La maggior parte degli organismi inseriti nel superphylum Lophotrochozoa possiede un apparato di alimentazione lofoforico o una larva trocoforica (da cui il nome contratto, "lopho-trocho-zoa"). Il lofoforo è una struttura di alimentazione composta da un insieme di tentacoli ciliati che circondano la bocca. Un trocoforo è una larva che nuota liberamente caratterizzata da due bande di ciglia che circondano un corpo a forma di cima. Ad alcuni dei phyla classificati come Lophotrochozoa potrebbe mancare una o entrambe queste strutture definitorie. Tuttavia, il loro posizionamento con i Lophotrochozoa è confermato quando vengono confrontati l'RNA ribosomiale e altre sequenze geniche. La sistematica di questo gruppo complesso non è ancora chiara e c'è ancora molto lavoro da fare per risolvere le relazioni cladistiche tra di loro.
Phylum dei Platelminti
modificaI vermi piatti sono organismi acelomati che includono molte forme libere e parassite. I vermi piatti non possiedono né larve lofofore né trocofore, sebbene le larve di un gruppo di vermi piatti, i Polycladida (chiamati così per il loro tratto digerente ramificato), siano considerate omologhe alle larve trocofore. La scissione a spirale è osservata anche nei polycladidi e in altri gruppi basali di vermi piatti. Il modello di sviluppo di alcune delle forme libere è oscurato da un fenomeno chiamato " anarchia dei blastomeri ", in cui si forma una sorta di larva di alimentazione temporanea, seguita da un raggruppamento di cellule all'interno dell'embrione che dà origine a un embrione di secondo stadio. Tuttavia, sia la monofilia dei vermi piatti che il loro posizionamento nei Lophotrochozoa sono stati supportati da analisi molecolari.
I Platelminti sono costituiti da due linee monofiletiche: i Catenulida e i Rhabditophora. I Catenulida, o "vermi a catena", sono un piccolo clade di poco più di 100 specie. Questi vermi si riproducono tipicamente asessualmente per gemmazione. Tuttavia, la prole non si stacca completamente dai genitori e la formazione assomiglia a una catena nell'aspetto. Tutti i vermi piatti qui discussi fanno parte dei Rhabditophora ("portatori di rabditi"). I rabditi sono strutture a forma di bastoncino scaricate nel muco prodotto da alcuni vermi piatti che vivono liberi; probabilmente servono sia per la difesa che per fornire trazione per lo scorrimento ciliare lungo il substrato. A differenza dei vermi piatti che vivono liberi, molte specie di trematodi e cestodi sono parassiti, compresi importanti parassiti degli esseri umani.
| a | b |
|---|---|
 |
 |
| I vermi piatti presentano una diversità significativa. (a) Un verme piatto blu Pseudoceros (Pseudoceros bifurcus) ; (b) un verme piatto maculato d'oro (Thysanozoon nigropapillosum) | |
I vermi piatti hanno tre strati di tessuto embrionale che danno origine ai tessuti epidermici (dall'ectoderma), al rivestimento del sistema digerente (dall'endoderma) e ad altri tessuti interni (dal mesoderma). Il tessuto epidermico è un singolo strato di cellule o uno strato di cellule fuse ( sincizio ) che ricopre due strati di muscolo, uno circolare e l'altro longitudinale. I tessuti mesodermici includono cellule mesenchimali che contengono collagene e supportano cellule secretorie che producono muco e altri materiali in superficie. Poiché i vermi piatti sono acelomati, lo strato mesodermico forma una massa solida tra la superficie epidermica esterna e la cavità del sistema digerente.
Processi fisiologici dei vermi piatti
modificaLe specie libere di vermi piatti sono predatori o spazzini. Le forme parassite si nutrono assorbendo i nutrienti forniti dai loro ospiti. La maggior parte dei vermi piatti, come la planaria mostrata nella Figura sotto, ha una cavità gastrovascolare ramificata piuttosto che un apparato digerente completo. In tali animali, la "bocca" viene anche utilizzata per espellere i materiali di scarto dall'apparato digerente e quindi funge anche da ano. (Alcune specie possono avere un secondo poro o apertura anale.) L'intestino può essere un semplice sacco o altamente ramificato. La digestione è principalmente extracellulare, con i materiali digeriti assorbiti nelle cellule del rivestimento intestinale tramite fagocitosi. Un gruppo di parassiti, i cestodi (cestodi), è completamente privo di un apparato digerente e assorbe il cibo digerito dall'ospite.
I vermi piatti hanno un sistema escretore con una rete di tubuli attaccati alle cellule a fiamma , le cui ciglia battono per dirigere i fluidi di scarto concentrati nei tubuli fuori dal corpo attraverso i pori escretori. Il sistema è responsabile della regolazione dei sali disciolti e dell'escrezione di rifiuti azotati. Il sistema nervoso è costituito da una coppia di cordoni nervosi laterali che corrono per tutta la lunghezza del corpo con connessioni trasversali tra loro. Due grandi gangli cerebrali, concentrazioni di corpi cellulari nervosi all'estremità anteriore del verme, sono associati a cellule fotosensoriali e chemiosensoriali. Non esiste né un sistema circolatorio né respiratorio, con lo scambio di gas e nutrienti dipendente dalla diffusione e dalle giunzioni cellula-cellula. Ciò limita necessariamente lo spessore del corpo in questi organismi, costringendoli a essere vermi "piatti". La maggior parte delle specie di vermi piatti è monoica (sia gli organi riproduttivi maschili che quelli femminili si trovano nello stesso individuo) e la fecondazione è tipicamente interna. La riproduzione asessuata per fissione è comune in alcuni gruppi.


Diversità dei vermi piatti
modificaI vermi piatti sono stati tradizionalmente divisi in quattro classi: Turbellaria, Monogenea, Trematoda e Cestoda ( Figura sotto). Tuttavia, le relazioni tra i membri di queste classi sono state recentemente rivalutate, con i turbellari in particolare ora considerati parafiletici, poiché i suoi discendenti possono anche includere membri delle altre tre classi. I membri del clade o classe Rhabditophora sono ora dispersi tra più ordini di Platyhelminthes, il più familiare dei quali è il Polycladida, che contiene i grandi vermi piatti marini; il Tricladida (che include Dugesia ["planaria"] e Planaria e i suoi parenti); e i principali ordini parassiti: Monogenea (ectoparassiti dei pesci), Trematoda (trematodi) e Cestoda (tenie), che insieme formano un clade monofiletico.
| a | b |
|---|---|
 |
 |
| c | d |
 |
 |
| Classi tradizionali di vermi piatti. Il phylum Platyhelminthes era precedentemente suddiviso in quattro classi. (a) La classe Turbellaria include il verme piatto policladide di Bedford ( Pseudobiceros bedfordi ), che è lungo circa 8-10 cm. (b) La classe parassitaria Monogenea include Dactylogyrus spp., comunemente chiamati trematodi branchiali, che sono lunghi circa 0,2 mm e hanno due ancore, indicate dalle frecce utilizzate per attaccare il parassita alle branchie del pesce ospite. (c) La classe Cestoda include tenie come questa Taenia saginata , che infetta sia i bovini che gli esseri umani e può raggiungere i 4-10 metri di lunghezza; l'esemplare mostrato qui è lungo circa quattro metri. (d) La classe Trematoda include Fascioloides magna (a destra) e Fasciola hepatica (due esemplari a sinistra, noti anche come trematodi epatici comuni). | |
La maggior parte dei vermi piatti che vivono liberi sono policladidi marini, sebbene le specie di tricladidi vivano in acqua dolce o in ambienti terrestri umidi, e vi siano diversi membri di altri ordini in entrambi gli ambienti. L'epidermide ventrale dei vermi piatti che vivono liberi è ciliata, il che ne facilita la locomozione. Alcuni vermi piatti che vivono liberi sono capaci di notevoli imprese di rigenerazione in cui un individuo può far ricrescere la testa o la coda dopo essere stato reciso, o persino diverse teste se la planaria viene tagliata longitudinalmente.
I monogenei sono ectoparassiti , per lo più di pesci, con cicli di vita semplici che consistono in una larva che nuota liberamente e che si attacca a un pesce, prima della sua trasformazione nella forma adulta ectoparassita. Il parassita ha un solo ospite e quell'ospite è solitamente molto specifico. I vermi possono produrre enzimi che digeriscono i tessuti dell'ospite, oppure possono semplicemente brucare muco superficiale e particelle di pelle. La maggior parte dei monogenei è ermafrodita, ma i gameti maschili si sviluppano prima e quindi la fecondazione incrociata è piuttosto comune.
I trematodi, o trematodi, sono parassiti interni di molluschi e molti altri gruppi, tra cui gli esseri umani. I trematodi hanno cicli di vita complessi che coinvolgono un ospite primario in cui avviene la riproduzione sessuale e uno o più ospiti secondari in cui avviene la riproduzione asessuata. L'ospite primario è solitamente un vertebrato e l'ospite secondario è quasi sempre un mollusco, in cui vengono prodotte più larve asessualmente. I trematodi, che si attaccano internamente all'ospite tramite una ventosa orale e mediale, sono responsabili di gravi malattie umane tra cui la schistosomiasi, causata da diverse specie di trematodi del sangue, Schistosoma spp. Varie forme di schistosomiasi infettano circa 200 milioni di persone ai tropici, causando danni agli organi, infezioni secondarie da batteri e sintomi cronici come la stanchezza. L'infezione si verifica quando l'uomo entra in acqua e le larve di metacercaria, rilasciate dall'ospite lumaca, localizzano e penetrano la pelle. Il parassita infetta vari organi del corpo e si nutre di globuli rossi prima di riprodursi.
Molte delle uova vengono rilasciate nelle feci e finiscono in un corso d'acqua, dove sono in grado di reinfettare l'ospite lumaca. Le uova, che hanno una punta, possono danneggiare il sistema vascolare dell'ospite umano, causando ulcerazioni, ascessi e diarrea sanguinolenta, ovunque risiedano, consentendo così ad altri patogeni di causare infezioni secondarie. Infatti, sono le uova del parassita a produrre la maggior parte dei principali effetti negativi della schistosomiasi. Molte uova non attraversano le vene dell'ospite per l'eliminazione e vengono spazzate via dal flusso sanguigno verso il fegato e altre sedi, dove possono causare una grave infiammazione. Nel fegato, le uova vaganti possono ostacolare la circolazione e causare cirrosi. Il controllo è difficile nelle aree povere in condizioni antigieniche e affollate e la prognosi è scarsa nelle persone con gravi infezioni da Schistosoma japonicum , senza un trattamento precoce.
I cestodi, o tenie, sono anche parassiti interni, principalmente di vertebrati ( Figura sotto). Le tenie, come quelle di Taenia spp, vivono nel tratto intestinale dell'ospite primario e rimangono fissate tramite una ventosa o uncini sull'estremità anteriore, o scolice, del corpo della tenia, che è essenzialmente una colonia di subunità simili chiamate proglottidi. Ogni proglottide può contenere un sistema escretore con cellule a fiamma, insieme a strutture riproduttive, sia maschili che femminili. Poiché sono così lunghe e piatte, le tenie non hanno bisogno di un sistema digerente; invece, assorbono i nutrienti dalla materia alimentare che le circonda nell'intestino dell'ospite per diffusione.
Le proglottidi vengono prodotte nello scolice e migrano gradualmente verso l'estremità della tenia; a questo punto, sono "mature" e tutte le strutture, eccetto le uova fecondate, sono degenerate. La maggior parte della riproduzione avviene tramite fecondazione incrociata tra diversi vermi nello stesso ospite, ma può verificarsi anche tra proglottidi. Le proglottidi mature si staccano dal corpo del verme e vengono rilasciate nelle feci dell'organismo. Le uova vengono mangiate da un ospite intermedio, in genere un altro vertebrato. Il verme giovane infetta l'ospite intermedio e prende residenza, di solito nel tessuto muscolare. Quando il tessuto muscolare viene consumato dall'ospite primario, il ciclo è completato. Esistono diversi parassiti della tenia degli esseri umani che vengono trasmessi mangiando carne di maiale, manzo o pesce crudi o poco cotti.

Phylum Rotifera
modificaI rotiferi ("wheel-bearer") appartengono a un gruppo di animali microscopici (circa 100 µm a 2 mm) per lo più acquatici che prendono il nome dalla corona , una coppia di strutture di alimentazione ciliate che sembrano ruotare se osservate al microscopio ottico ( Figura sotto). Sebbene il loro stato tassonomico sia attualmente in evoluzione, un trattamento colloca i rotiferi in tre classi: Bdelloidea, Monogononta e Seisonidea. Inoltre, i parassiti "vermi dalla testa spinosa" attualmente nel phylum Acanthocephala, sembrano essere rotiferi modificati e saranno probabilmente inseriti nel gruppo nel prossimo futuro. Indubbiamente i rotiferi continueranno a essere rivisti man mano che saranno disponibili maggiori prove filogenetiche.
Il corpo pseudocelomato di un rotiferi è notevolmente complesso per un animale così piccolo (più o meno delle dimensioni di un Paramecium ) ed è diviso in tre sezioni: una testa (che contiene la corona), un tronco (che contiene la maggior parte degli organi interni) e il piede . Una cuticola, rigida in alcune specie e flessibile in altre, ricopre la superficie del corpo. Hanno sia muscoli scheletrici associati alla locomozione sia muscoli viscerali associati all'intestino, entrambi composti da singole cellule. I rotiferi sono in genere organismi che nuotano liberamente o planctonici (alla deriva), ma le dita o le estensioni del piede possono secernere un materiale appiccicoso per aiutarli ad aderire alle superfici. La testa contiene una serie di macchie oculari e un "cervello" bilobato, con nervi che si estendono nel corpo.
| a | b |
|---|---|
 |
 |
| Rotiferi. Sono mostrati esempi di due delle tre classi di rotiferi. (a) Le specie della classe Bdelloidea sono caratterizzate da una grande corona. Gli animali interi al centro di questa micrografia elettronica a scansione sono mostrati circondati da diversi set di mascelle dal mastax dei rotiferi. (b) Polyarthra , della più grande classe di rotiferi Monogononta, ha una corona più piccola dei rotiferi bdelloidi e una singola gonade, che dà il nome alla classe. | |
I rotiferi si trovano comunemente in acqua dolce e in alcuni ambienti di acqua salata in tutto il mondo. Come filtratori, mangiano materiale morto, alghe e altri organismi viventi microscopici e sono quindi componenti molto importanti delle reti alimentari acquatiche. Il cibo di un rotiferi è diretto verso la bocca dalla corrente creata dal movimento delle ciglia coronali. Le particelle di cibo entrano nella bocca e viaggiano prima verso il mastax , una faringe muscolare con strutture simili a mascelle dentate. Esempi delle mascelle di vari rotiferi sono visibili nella Figura sotto a. Il cibo masticato passa vicino alle ghiandole digestive e salivari, nello stomaco e poi nell'intestino. I rifiuti digestivi ed escretori vengono raccolti in una vescica cloacale prima di essere rilasciati dall'ano.
Sono state identificate circa 2.200 specie di rotiferi. La figura sotto mostra l'anatomia di un rotifero appartenente alla classe Bdelloidea. Alcuni rotiferi sono organismi dioici e presentano dimorfismo sessuale (maschi e femmine hanno forme diverse). In molte specie dioiche, i maschi hanno una vita breve e sono più piccoli, senza apparato digerente e con un solo testicolo. Molte specie di rotiferi presentano aplodiploidia, un metodo di determinazione del sesso in cui un uovo fecondato si sviluppa in una femmina e un uovo non fecondato si sviluppa in un maschio. Tuttavia, la riproduzione nei rotiferi bdelloidei è esclusivamente partenogenetica e sembra esserlo da milioni di anni: quindi, tutti i rotiferi bdelloidi e la loro progenie sono femmine! I bdelloidei possono compensare questa insularità genetica prendendo in prestito geni dal DNA di altre specie. Fino al 10% del genoma di un bdelloideo comprende geni importati da specie correlate. Alcune uova di rotiferi possono restare in uno stato di dormienza prolungato per proteggersi dalle condizioni ambientali difficili.

Phylum Nemertea
modificaI Nemertea sono colloquialmente noti come vermi a nastro o vermi proboscide. La maggior parte delle specie del phylum Nemertea sono marine e prevalentemente bentoniche (abitanti dei fondali), con una stima di 900 specie note. Tuttavia, i nemerteani sono stati registrati anche in habitat terrestri di acqua dolce e molto umidi. La maggior parte dei nemerteani sono carnivori e si nutrono di vermi, vongole e crostacei. Alcune specie sono spazzini e alcune, come Malacobdella grossa , hanno anche sviluppato relazioni commensali con i molluschi. Le specie economicamente importanti hanno a volte devastato la pesca commerciale di vongole e granchi. I nemerteani non hanno quasi predatori e due specie sono vendute come esche per pesci.
Morfologia
modificaI Nemertea variano in dimensioni da 1 cm a diversi metri. Mostrano simmetria bilaterale e notevoli proprietà contrattili. Grazie alla loro contrattilità, possono cambiare la loro presentazione morfologica in risposta a stimoli ambientali. Gli animali del phylum Nemertea sono animali morbidi e non segmentati, con una morfologia simile a un tubo appiattito. ( Figura sotto).

Una caratteristica unica di questo phylum è la presenza di una proboscide eversibile racchiusa in una tasca chiamata rincocele (non parte del celoma effettivo dell'animale). La proboscide è situata dorsalmente all'intestino e funge da arpione o tentacolo per la cattura del cibo. In alcune specie è ornata di barbigli. Il rincocele è una cavità piena di liquido che si estende dalla testa fino a quasi due terzi della lunghezza dell'intestino in questi animali ( Figura sotto). La proboscide può essere estesa dalla pressione idrostatica generata dalla contrazione del muscolo del rincocele e retratta da un muscolo retrattore attaccato alla parete posteriore del rincocele.

Sistema digerente
modificaI nemertei, che sono principalmente predatori di anellidi e crostacei, hanno un apparato digerente ben sviluppato. Un'apertura della bocca ventrale al rincocele conduce all'intestino anteriore, seguito dall'intestino. L'intestino è presente sotto forma di tasche diverticolari e termina in un retto che si apre tramite un ano. Le gonadi sono intervallate dalle tasche diverticolari intestinali e si aprono verso l'esterno tramite pori genitali.
I nemertei sono talvolta classificati come acoel, ma poiché il loro sistema circolatorio chiuso deriva dalla cavità celomatica dell'embrione, possono essere considerati celomici. Il loro sistema circolatorio è costituito da un anello chiuso formato da una coppia connessa di vasi sanguigni laterali. Alcune specie possono anche avere un vaso dorsale o vasi trasversali oltre a quelli laterali. Sebbene il fluido circolatorio contenga cellule, è spesso incolore. Tuttavia, le cellule del sangue di alcune specie contengono emoglobina e altri pigmenti gialli o verdi. I vasi sanguigni sono contrattili, sebbene di solito non vi sia un percorso circolatorio regolare, e il movimento del sangue è anche facilitato dalla contrazione dei muscoli nella parete corporea. La circolazione dei fluidi nel rincocele è più o meno indipendente dalla circolazione sanguigna, sebbene rami ciechi dai vasi sanguigni nella parete del rincocele possano mediare lo scambio di materiali tra di loro. Una coppia di protonefridi , o tubuli escretori, è presente in questi animali per facilitare l'osmoregolazione. Lo scambio gassoso avviene attraverso la pelle.
Sistema nervoso
modificaI nemerteani hanno un "cervello" composto da quattro gangli situati all'estremità anteriore, attorno al rincocele. Cordoni nervosi longitudinali appaiati emergono dai gangli cerebrali e si estendono fino all'estremità posteriore. In alcune specie si trovano cordoni nervosi aggiuntivi. È interessante notare che il cervello può contenere emoglobina, che funge da riserva di ossigeno. Gli ocelli o macchie oculari sono presenti in coppie, in multipli di due nella parte anteriore del corpo. Si ipotizza che le macchie oculari abbiano origine dal tessuto neurale e non dall'epidermide.
Riproduzione
modificaI Nemertea, come i vermi piatti, hanno eccellenti capacità di rigenerazione e in alcune specie si osserva la riproduzione asessuata per frammentazione. La maggior parte degli animali del phylum Nemertea è dioica, sebbene le specie di acqua dolce possano essere ermafrodite. Le cellule staminali che diventano gameti si aggregano all'interno delle gonadi poste lungo il tratto digerente. Le uova e lo sperma vengono rilasciati nell'acqua e la fecondazione avviene esternamente. Come la maggior parte dei protostomi lofotrocozoi, la scissione è a spirale e lo sviluppo è solitamente diretto, sebbene alcune specie abbiano una larva simile a un trocoforo, in cui un giovane verme è costruito da una serie di dischi immaginali che iniziano come invaginazioni dalla superficie corporea della larva.
Superphylum Lophotrochozoa: Molluschi e Anellidi
modificaGli anellidi e i molluschi sono i più noti tra i protostomi dei lofotrocozoi. Sono anche i lofotrocozoi più "tipici", poiché entrambi i gruppi includono specie acquatiche con larve trocofore, che uniscono entrambi i taxa in una comune discendenza. Questi phylum mostrano come un piano corporeo flessibile possa portare al successo biologico in termini di abbondanza e diversità di specie. Il phylum Mollusca ha il secondo maggior numero di specie di tutti i phylum animali con quasi 100.000 specie esistenti descritte e circa 80.000 specie estinte descritte. Infatti, si stima che circa il 25 percento di tutte le specie marine conosciute siano molluschi! Gli anellidi e i molluschi sono entrambi eucoelomati bilateralmente simmetrici, cefalizzati, triploblastici, schizoceli Includono animali che probabilmente vedrai nel tuo cortile o nel tuo piatto!
Phylum Mollusca
modificaIl nome "Mollusca" significa corpo "morbido", poiché le prime descrizioni di molluschi derivano da osservazioni di seppie "mollicce" e senza guscio. I molluschi sono prevalentemente un gruppo di animali marini; tuttavia, sono noti anche per abitare habitat di acqua dolce e terrestri. Questo enorme phylum comprende chitoni, gusci di zanne, lumache, lumache di mare, nudibranchi, farfalle di mare, vongole, cozze, ostriche, calamari, polpi e nautilus. I molluschi mostrano un'ampia gamma di morfologie in ogni classe e sottoclasse, ma condividono alcune caratteristiche chiave ( Figura sotto). La principale struttura locomotoria è solitamente un piede muscolare . La maggior parte degli organi interni è contenuta in una regione chiamata massa viscerale . Sovrastante la massa viscerale c'è una piega di tessuto chiamata mantello ; all'interno della cavità formata dal mantello si trovano strutture respiratorie chiamate branchie , che in genere si ripiegano sulla massa viscerale. Le bocche della maggior parte dei molluschi, eccetto i bivalvi (ad esempio, le vongole) contengono un organo di alimentazione specializzato chiamato radula , una struttura abrasiva simile a una lingua. Infine, il mantello secerne una conchiglia indurita dal carbonato di calcio nella maggior parte dei molluschi, sebbene questa sia notevolmente ridotta nella classe dei cefalopodi, che contiene i polpi e i calamari.


Esercizio: quale delle seguenti affermazioni sull'anatomia di un mollusco è falsa?
- La maggior parte dei molluschi è dotata di una radula per triturare il cibo.
- Una ghiandola digestiva è collegata allo stomaco.
- Il tessuto sottostante la conchiglia è chiamato mantello.
- L'apparato digerente è composto da un ventriglio, uno stomaco, una ghiandola digestiva e l'intestino.
Il piede muscoloso è l'organo più ventrale, mentre il mantello è l'organo dorsale limitante che si ripiega sulla massa viscerale . Il piede, che viene utilizzato per la locomozione e l'ancoraggio, varia in forma e funzione, a seconda del tipo di mollusco in esame. Nei molluschi con conchiglia, il piede è solitamente delle stesse dimensioni dell'apertura della conchiglia. Il piede è sia retrattile che estensibile. Nella classe Cefalopodi ("testa-piede"), il piede assume la forma di un imbuto per espellere l'acqua ad alta velocità dalla cavità del mantello; e il margine anteriore del piede è stato modificato in un cerchio di braccia e tentacoli .
La massa viscerale è presente sopra il piede, nella gobba viscerale. Questa massa contiene i sistemi digerente, nervoso, escretore, riproduttivo e respiratorio. Le specie di molluschi che sono esclusivamente acquatiche hanno branchie che si estendono nella cavità del mantello, mentre alcune specie terrestri hanno "polmoni" formati dal rivestimento della cavità del mantello. I molluschi sono eucoelomati schizoceli, ma la cavità celomatica negli animali adulti è stata ampiamente ridotta a una cavità attorno al cuore. Tuttavia, un celoma ridotto a volte circonda anche le gonadi, parte dei reni e l'intestino. Questa riduzione celomatica complessiva rende la cavità del mantello la principale camera interna del corpo.
La maggior parte dei molluschi ha uno speciale organo a forma di raspa, la radula , che porta denti chitinosi simili a lima. La radula è presente in tutti i gruppi, eccetto i bivalvi, e serve a sminuzzare o raschiare il cibo prima che entri nel tratto digerente. Il mantello (noto anche come pallio ) è l'epidermide dorsale nei molluschi; tutti i molluschi, eccetto alcuni cefalopodi, sono specializzati nel secernere un guscio calcareo che protegge il corpo molle dell'animale.
La maggior parte dei molluschi sono animali dioici e la fecondazione avviene esternamente, sebbene ciò non avvenga nei molluschi terrestri, come lumache e limacce, o nei cefalopodi. Nella maggior parte dei molluschi acquatici, lo zigote si schiude e produce una larva trocofora , con diverse fasce di ciglia attorno a un corpo a forma di cima e un ulteriore ciuffo apicale di ciglia. In alcune specie, la trocofora può essere seguita da ulteriori stadi larvali, come una larva veliger , prima della metamorfosi finale nella forma adulta. La maggior parte dei cefalopodi si sviluppa direttamente in piccole versioni della loro forma adulta.
Classificazione del phylum Mollusca
modificaIl phylum Mollusca comprende un gruppo molto diversificato di organismi che esibisce una varietà drammatica di forme, che vanno dai chitoni alle lumache ai calamari, questi ultimi dei quali mostrano tipicamente un alto grado di intelligenza. Questa variabilità è una conseguenza della modifica delle regioni corporee di base, in particolare il piede e il mantello. Il phylum è organizzato in otto classi: Caudofoveata, Solenogastres, Monoplacophora, Polyplacophora, Gastropoda, Cephalopoda, Bivalvia e Scaphopoda. Sebbene ogni classe di molluschi sembri essere monofiletica, la loro relazione reciproca non è chiara e ancora in fase di revisione.
Sia i Caudofoveata che i Solenogastres includono animali senza conchiglia, simili a vermi, che si trovano principalmente in habitat marini bentonici. Sebbene questi animali non abbiano una conchiglia calcarea, ottengono una certa protezione da spicole calcaree incorporate in una cuticola che ricopre la loro epidermide. La cavità del mantello è ridotta ed entrambi i gruppi sono privi di occhi, tentacoli e nefridi (organi escretori). I Caudofoveata possiedono una radula, ma i Solenogastres non hanno né radula né branchie. Anche il piede è ridotto nei Solenogastres e assente nei Caudofoveata.
A lungo ritenuto estinto, il primo esemplare vivente di Monoplacophora, Neopilina galatheae , fu scoperto nel 1952 sul fondale oceanico vicino alla costa occidentale della Costa Rica. Oggi sono state descritte oltre 25 specie. I membri della classe Monoplacophora ("con una piastra") possiedono un singolo guscio a forma di calotta che ricopre il corpo dorsale. La morfologia del guscio e dell'animale sottostante può variare da circolare a ovata. Hanno una radula semplice, un sistema digerente ad anello, più paia di organi escretori e un paio di gonadi. Tra il piede e il bordo del mantello si trovano più branchie.
Gli animali della classe Polyplacophora ("che portano molte piastre") sono comunemente noti come "chitoni" e portano otto piastre calcaree che compongono la conchiglia dorsale ( Figura sotto). Questi animali hanno un piede ventrale largo che è adattato per l'aspirazione su rocce e altri substrati e un mantello che si estende oltre il bordo della conchiglia. Le spine calcaree sul bordo esposto del mantello forniscono protezione dai predatori. La respirazione è facilitata da più paia di branchie nella cavità del mantello. Il sangue dalle branchie viene raccolto in un cuore posteriore e quindi inviato al resto del corpo in un emocele , un sistema di circolazione aperto in cui il sangue è contenuto in camere collegate che circondano vari organi anziché all'interno di singoli vasi sanguigni. La radula, che ha denti composti da una magnetite ultradura, viene utilizzata per raschiare gli organismi alimentari dalle superfici rocciose. È stato dimostrato che i denti di chitone presentano la maggiore durezza e rigidità di qualsiasi altro materiale biominerale finora riportato, essendo fino a tre volte più duri dello smalto umano e dei gusci dei molluschi a base di carbonato di calcio.
Il sistema nervoso è rudimentale, con solo gangli buccali o "delle guance" presenti all'estremità anteriore. Molteplici piccole strutture sensoriali, tra cui fotosensori, si estendono dal mantello in canali nello strato superiore della conchiglia. Queste strutture sono chiamate esteti e sono esclusive dei chitoni. Un'altra struttura sensoriale sotto la radula è utilizzata per campionare l'ambiente di alimentazione. Un singolo paio di nefridi è utilizzato per l'escrezione di rifiuti azotati.

La classe Bivalvia ("due-valve") include vongole, ostriche, cozze, capesante, geoduck e teredini. Alcuni bivalvi sono quasi microscopici, mentre altri, nel genere Tridacna, possono essere lunghi un metro e pesare 225 chilogrammi. I membri di questa classe si trovano sia in habitat marini che di acqua dolce. Come suggerisce il nome, i bivalvi sono racchiusi in valve o gusci in due parti ( Figura sotto a) fuse sul lato dorsale da legamenti a cerniera e denti del guscio sul lato ventrale che mantengono le due metà allineate. I due gusci, che consistono in uno strato organico esterno, uno strato prismatico intermedio e uno strato madreperlaceo molto liscio, sono uniti nella parte più vecchia del guscio chiamata umbone. I muscoli adduttori e abduttori anteriori e posteriori chiudono e aprono rispettivamente il guscio.
Il corpo complessivo del bivalve è appiattito lateralmente; il piede è a forma di cuneo; e la regione della testa è poco sviluppata (senza una bocca evidente). I bivalvi sono filtratori e in questa classe di molluschi è assente una radula. La cavità del mantello è fusa lungo i bordi, fatta eccezione per le aperture per il piede e per l'assunzione e l'espulsione dell'acqua, che viene fatta circolare attraverso la cavità del mantello dalle azioni dei sifoni di ingresso e di uscita. Durante l'assunzione di acqua da parte del sifone di ingresso, le particelle di cibo vengono catturate dalle branchie posteriori accoppiate (ctenidi) e quindi trasportate dal movimento delle ciglia in avanti verso la bocca. L'escrezione e l'osmoregolazione sono eseguite da una coppia di nefridi. Ocelli e altre strutture sensoriali sono situati lungo il bordo del mantello in alcune specie. Gli "occhi" sono particolarmente evidenti nelle capesante ( Figura sotto b). Tre paia di gangli collegati regolano l'attività di diverse strutture corporee.
| a | b |
|---|---|
 |
 |
| Bivalvi. Questi mitili (a), trovati nella zona intertidale in Cornovaglia, Inghilterra, mostrano la conchiglia bivalve. La capasanta Argopecten irradians (b) ha una conchiglia scanalata e vistosi ocelli | |
Una delle funzioni del mantello è quella di secernere la conchiglia. Alcuni bivalvi, come le ostriche e le cozze, possiedono la capacità unica di secernere e depositare una madreperla calcarea o "madreperla" attorno a particelle estranee che potrebbero entrare nella cavità del mantello. Questa proprietà è stata sfruttata commercialmente per produrre perle.
Collegamento all'apprendimento
modificaGuarda le animazioni dell'alimentazione dei bivalvi: osserva il processo di alimentazione di vongole e cozze in questi siti.
Più della metà delle specie di molluschi appartengono alla classe Gastropoda ("piede di stomaco"), che include molluschi ben noti come lumache, limacce, conchiglie, cipree, patelle e buccini. I gasteropodi acquatici includono sia specie marine che d'acqua dolce e tutti i molluschi terrestri sono gasteropodi. I gasteropodi includono specie con conchiglia e specie senza conchiglia. I corpi dei gasteropodi sono asimmetrici e solitamente presentano una conchiglia arrotolata ( Figura sotto a). Le conchiglie possono essere planospirali (come un tubo da giardino arrotolato), comunemente osservate nelle lumache da giardino, o conispirali (come una scala a chiocciola), comunemente osservate nelle conchiglie marine. Le conchiglie di ciprea hanno una superficie levigata perché il mantello si estende sopra la parte superiore della conchiglia mentre viene secreto.
| a | b |
|---|---|
 |
 |
| Gasteropodi. Le lumache (a) e le chiocciole (b) sono entrambe gasteropodi, ma le lumache non hanno conchiglia | |
Una caratteristica fondamentale di alcuni gasteropodi è lo sviluppo embrionale della torsione . Durante questo processo, il mantello e la massa viscerale vengono ruotati attorno all'asse perpendicolare al centro del piede per portare l'apertura anale in avanti appena dietro la testa ( Figura sotto), creando una situazione molto particolare. La branchia sinistra, il rene e l'atrio cardiaco sono ora sul lato destro, mentre la branchia destra originale, il rene e l'atrio cardiaco sono sul lato sinistro. Ancora più strano, i cordoni nervosi sono stati attorcigliati e contorti in uno schema a otto. Grazie allo spazio reso disponibile dalla torsione nella cavità del mantello, l'estremità sensibile della testa dell'animale può ora essere ritirata nella protezione del guscio e il piede più resistente (e talvolta la copertura protettiva o l'opercolo ) forma una barriera verso l'esterno. La strana disposizione che risulta dalla torsione pone un serio problema igienico-sanitario creando la possibilità che i rifiuti vengano rilavati sulle branchie, causando sporcizia. In realtà non esiste una spiegazione davvero perfetta per lo sviluppo embrionale della torsione, e si sa che alcuni gruppi che in passato presentavano la torsione nei loro gruppi ancestrali hanno ora invertito il processo.
I gasteropodi hanno anche un piede modificato per strisciare. La maggior parte dei gasteropodi ha una testa ben definita con tentacoli e occhi. Una radula complessa viene utilizzata per raschiare le particelle di cibo. Nei gasteropodi acquatici, la cavità del mantello racchiude le branchie (ctenidi), ma nei gasteropodi terrestri, il mantello stesso è la principale struttura respiratoria, che agisce come una specie di polmone. I nefridi ("reni") si trovano anche nella cavità del mantello.

Il veleno di lumaca può essere utilizzato come antidolorifico farmacologico?
modificaLe lumache marine del genere Conus ( Figura sotto) attaccano le prede con un pungiglione velenoso, modificato dalla radula. La tossina rilasciata, nota come conotossina, è un peptide con legami disolfuro interni. Le conotossine possono causare paralisi negli esseri umani, il che indica che questa tossina attacca i bersagli neurologici. È stato dimostrato che alcune conotossine bloccano i canali ionici neuronali. Queste scoperte hanno portato i ricercatori a studiare le conotossine per possibili applicazioni mediche.
Le conotossine sono un'area entusiasmante di potenziale sviluppo farmacologico, poiché questi peptidi possono essere eventualmente modificati e utilizzati in specifiche condizioni mediche per inibire l'attività di specifici neuroni. Ad esempio, le conotossine o le loro modifiche possono essere utilizzate per indurre la paralisi nei muscoli in specifiche applicazioni sanitarie, simili all'uso della tossina botulinica. Poiché l'intero spettro delle conotossine, così come i loro meccanismi d'azione, non è completamente noto, lo studio delle loro potenziali applicazioni è ancora agli inizi. La maggior parte delle ricerche fino ad oggi si è concentrata sul loro utilizzo per trattare malattie neurologiche. Hanno anche mostrato una certa efficacia nell'alleviare il dolore cronico e il dolore associato a condizioni come la sciatica e l'herpes zoster. Lo studio e l'uso delle biotossine, tossine derivate da organismi viventi, sono un esempio eccellente dell'applicazione della scienza biologica alla medicina moderna.
| a | b |
|---|---|
 |
 |
| Conus. I membri del genere Conus producono neurotossine che un giorno potrebbero avere usi medici. Il tubo sopra gli occhi è un sifone usato sia per far circolare l'acqua sulle branchie sia per campionare l'acqua per prove chimiche di prede nelle vicinanze. Notare gli occhi sotto il sifone. La proboscide, attraverso la quale viene proiettato l'arpione velenoso, si trova tra gli occhi. | |
Classe Cefalopodi (animali con "testa e piede"), include polpi, calamari, seppie e nautilus. I cefalopodi includono sia animali con conchiglia che animali in cui la conchiglia è ridotta o assente. Nel Nautilus con conchiglia, la conchiglia a spirale è multicamera. Queste camere sono riempite di gas o acqua per regolare la galleggiabilità. Un sifone attraversa le camere ed è questo tubo che regola la quantità di acqua e gas (miscela di azoto, anidride carbonica e ossigeno) presenti nelle camere. Ammoniti e altre conchiglie nautiloidi sono comunemente osservate nei reperti fossili. La struttura della conchiglia nei calamari e nelle seppie è ridotta ed è presente internamente sotto forma di penna di calamaro e osso di seppia, rispettivamente. L'osso di seppia è venduto nei negozi di animali per aiutare a lisciare i becchi degli uccelli e anche per fornire a uccelli come galline ovaiole e quaglie una fonte naturale poco costosa di carbonato di calcio. Esempi di cefalopodi sono mostrati nella Figura sotto.
I cefalopodi possono mostrare una colorazione vivida e in rapido cambiamento, quasi come insegne al neon lampeggianti. In genere queste esibizioni lampeggianti si vedono nei calamari e nei polpi, dove possono essere utilizzate per mimetizzarsi e forse come segnali per le esibizioni di accoppiamento. Dovremmo notare, tuttavia, che i ricercatori non sono del tutto sicuri se i calamari possano effettivamente vedere il colore, o se lo vedano allo stesso modo in cui lo vediamo noi. Sappiamo che i pigmenti nella pelle sono contenuti in speciali cellule pigmentarie ( cromatofori ), che possono espandersi o contrarsi per produrre diversi modelli di colore. Ma i cromatofori possono creare solo pigmentazione gialla, rossa, marrone e nera; tuttavia, sotto di loro c'è un set completamente diverso di elementi chiamati iridofori e leucofori che riflettono la luce e possono creare blu, verde e bianco. È possibile che la pelle del calamaro possa effettivamente essere in grado di rilevare un po' di luce da sola, senza nemmeno aver bisogno dei suoi occhi!
Tutti gli animali di questa classe sono predatori carnivori e hanno mascelle simili a becchi oltre alla radula. I cefalopodi includono i più intelligenti tra i molluschi e hanno un sistema nervoso ben sviluppato insieme a occhi che formano immagini. A differenza di altri molluschi, hanno un sistema circolatorio chiuso, in cui il sangue è interamente contenuto in vasi piuttosto che in un emocele.
Il piede è lobato e suddiviso in braccia e tentacoli. Su braccia e tentacoli di polpi e calamari sono presenti ventose con anelli chitinizzati. I sifoni sono ben sviluppati e l'espulsione dell'acqua è utilizzata come modalità primaria di locomozione, che assomiglia alla propulsione a getto. Le branchie (ctenidi) sono attaccate alla parete della cavità del mantello e sono servite da grandi vasi sanguigni, ciascuno con un proprio cuore. Una coppia di nefridi è presente all'interno della cavità del mantello per l'equilibrio idrico e l'escrezione di rifiuti azotati. I cefalopodi come calamari e polpi producono anche seppia o un inchiostro scuro, che contiene melanina. La ghiandola dell'inchiostro è situata tra le branchie e può essere rilasciata nel flusso d'acqua in uscita. Le nuvole di inchiostro possono essere utilizzate come "cortina fumogena" per nascondere l'animale dai predatori durante un rapido tentativo di fuga o per creare un'immagine falsa per distrarre i predatori.
I cefalopodi sono dioici. I membri di una specie si accoppiano e la femmina depone le uova in una nicchia appartata e protetta. Le femmine di alcune specie si prendono cura delle uova per un lungo periodo di tempo e potrebbero finire per morire durante quel periodo. Mentre la maggior parte degli altri molluschi acquatici produce larve trocofore, le uova dei cefalopodi si sviluppano direttamente in un giovane senza uno stadio larvale intermedio.
| a | b |
|---|---|
 |
 |
| c | d |
.jpg) |
 |
| Cefalopodi. (a) nautilus, (b) seppia gigante, (c) calamaro di barriera e (d) polpo dagli anelli blu sono tutti membri della classe Cefalopodi | |
I membri della classe Scaphopoda ("piedi di barca") sono conosciuti colloquialmente come "gusci di zanna" o "gusci di dente", come è evidente quando si esamina Dentalium , uno dei pochi generi di scafopodi rimasti ( Figura sotto). Gli scafopodi sono solitamente sepolti nella sabbia con l'apertura anteriore esposta all'acqua. Questi animali hanno un singolo guscio conico, aperto su entrambe le estremità. La testa non è ben sviluppata, ma la bocca, contenente una radula, si apre tra un gruppo di tentacoli che terminano in bulbi ciliati usati per catturare e manipolare le prede. Gli scafopodi hanno anche un piede simile a quello visto nei bivalvi. Gli ctenidi sono assenti in questi animali; la cavità del mantello forma un tubo aperto su entrambe le estremità e funge da struttura respiratoria in questi animali.

Phylum Anellidi
modificaIl phylum Annelida comprende i veri vermi segmentati. Questi animali si trovano in habitat marini, terrestri e di acqua dolce, ma la presenza di acqua o umidità è un fattore critico per la loro sopravvivenza negli habitat terrestri. Gli anellidi sono spesso chiamati "vermi segmentati" a causa della loro caratteristica chiave del metamerismo , o vera segmentazione. Circa 22.000 specie sono state descritte nel phylum Annelida, che include vermi policheti (anellidi marini con appendici multiple) e oligocheti (lombrichi e sanguisughe). Alcuni animali in questo phylum mostrano simbiosi parassite e commensali con altre specie nel loro habitat.
Morfologia
modificaGli anellidi mostrano simmetria bilaterale e sono simili a vermi nella morfologia complessiva. Il nome del phylum deriva dalla parola latina annulus , che significa piccolo anello, una descrizione appropriata della segmentazione ad anello del corpo. Gli anellidi hanno un piano corporeo con segmentazione metamerica, in cui diverse caratteristiche morfologiche interne ed esterne si ripetono in ogni segmento corporeo. Il metamerismo consente agli animali di diventare più grandi aggiungendo "compartimenti", rendendo al contempo più efficiente il loro movimento. Il corpo complessivo può essere suddiviso in testa, corpo e pigidio (o coda). Durante lo sviluppo, i segmenti dietro la testa sorgono in sequenza da una regione di crescita anteriore al pigidio, un modello chiamato crescita teloblastica . Negli Oligocheti, il clitello è una struttura riproduttiva che genera muco per facilitare il trasferimento dello sperma e produce anche un "bozzolo", all'interno del quale avviene la fecondazione; appare come una banda permanente e fusa situata sul terzo anteriore dell'animale ( Figura sotto).

Anatomia
modificaL'epidermide è protetta da una cuticola collagena esterna, che è molto più sottile della cuticola presente negli ecdisozoi e non richiede una periodica caduta per la crescita. Muscoli sia circolari che longitudinali sono situati all'interno dell'epidermide. Setole chitinose chiamate setole (o chete) sono ancorate all'epidermide, ciascuna con il proprio muscolo. Nei policheti, le setole sono portate su appendici appaiate chiamate parapodi .
La maggior parte degli anellidi ha un sistema digerente ben sviluppato e completo. I meccanismi di alimentazione variano ampiamente nel phylum. Alcuni policheti sono filtratori che usano appendici simili a piume per raccogliere piccoli organismi. Altri hanno tentacoli, mascelle o una faringe eversibile per catturare le prede. I lombrichi raccolgono piccoli organismi dal terreno mentre vi scavano dentro e la maggior parte delle sanguisughe sono ematofagi armati di denti o di una proboscide muscolare. Nei lombrichi, il tratto digerente comprende una bocca, una faringe muscolare, un esofago, un gozzo e un ventriglio muscolare. Il ventriglio conduce all'intestino, che termina in un'apertura anale nel segmento terminale. Una vista trasversale di un segmento corporeo di un lombrico è mostrata nella Figura 28.30 ; ogni segmento è limitato da un setto membranoso che divide la cavità celomatica in una serie di compartimenti.
La maggior parte degli anellidi possiede un sistema circolatorio chiuso di vasi sanguigni dorsali e ventrali che corrono parallelamente al canale alimentare, nonché capillari che servono i singoli tessuti. Inoltre, i vasi dorsali e ventrali sono collegati da anse trasversali in ogni segmento. Alcuni policheti e sanguisughe hanno un sistema aperto in cui i principali vasi sanguigni si aprono in un emocele. In molte specie, il sangue contiene emoglobina, ma non è contenuta nelle cellule. Gli anellidi non hanno un sistema respiratorio ben sviluppato e lo scambio di gas avviene attraverso la superficie corporea umida. Nei policheti, i parapodi sono altamente vascolarizzati e fungono da strutture respiratorie. L'escrezione è facilitata da una coppia di metanefridi (un tipo di "rene" primitivo costituito da un tubulo contorto e un imbuto aperto e ciliato) che è presente in ogni segmento verso il lato ventrale. Gli anellidi mostrano sistemi nervosi ben sviluppati con un anello di gangli fusi presenti attorno alla faringe. Il cordone nervoso è in posizione ventrale e presenta nodi o gangli ingrossati in ciascun segmento.


Gli anellidi possono essere monoici con gonadi permanenti (come nei lombrichi e nelle sanguisughe) o dioici con gonadi temporanee o stagionali (come nei policheti). Tuttavia, la fecondazione incrociata è preferita anche negli animali ermafroditi. I lombrichi possono mostrare una fecondazione reciproca simultanea quando sono allineati per la copulazione. Alcune sanguisughe cambiano sesso durante la loro vita riproduttiva. Nella maggior parte dei policheti, la fecondazione è esterna e lo sviluppo include una larva trocofora, che poi metamorfizza nella forma adulta. Negli oligocheti, la fecondazione è tipicamente interna e le uova fecondate si sviluppano in un bozzolo prodotto dal clitello; lo sviluppo è diretto. I policheti sono eccellenti rigeneratori e alcuni si riproducono persino asessualmente per gemmazione o frammentazione.
Video
modificaQuesto video offre uno sguardo ravvicinato all'anatomia degli anellidi.
Classificazione del phylum degli anellidi
modificaIl phylum Annelida contiene la classe Polychaeta (i policheti) e la classe Oligochaeta (i lombrichi, le sanguisughe e i loro parenti). I lombrichi e le sanguisughe formano un clade monofiletico all'interno dei policheti, che sono quindi parafiletici come gruppo.
Esistono più di 22.000 specie diverse di anellidi, e più della metà di queste sono policheti marini ("molte setole"). Nei policheti, le setole sono disposte in gruppi sui loro parapodi, appendici carnose, piatte e appaiate che sporgono da ogni segmento. Molti policheti usano i loro parapodi per strisciare sul fondale marino, ma altri sono adattati per nuotare o galleggiare. Alcuni sono sessili e vivono in tubi. Alcuni policheti vivono vicino alle sorgenti idrotermali. Questi vermi tubolari di acque profonde non hanno un tratto digerente, ma hanno una relazione simbiotica con i batteri che vivono nei loro corpi.
I lombrichi sono i membri più abbondanti della classe Oligochaeta ("poche setole"), distinti dalla presenza di un clitello permanente e dal piccolo numero di chete ridotte su ogni segmento. (Ricorda che gli oligocheti non hanno parapodi.) La sottoclasse degli oligocheti Hirudinea, include sanguisughe come la sanguisuga medicinale, Hirudo medicinalis , che è efficace nell'aumentare la circolazione sanguigna e rompere i coaguli di sangue, e quindi può essere utilizzata per trattare alcuni disturbi circolatori e malattie cardiovascolari. Il loro uso risale a migliaia di anni fa. Questi animali producono un clitello stagionale, a differenza del clitello permanente di altri oligocheti. Una differenza significativa tra le sanguisughe e gli altri anellidi è la mancanza di setole e lo sviluppo di ventose alle estremità anteriore e posteriore, che vengono utilizzate per attaccarsi all'animale ospite. Inoltre, nelle sanguisughe, la segmentazione della parete corporea potrebbe non corrispondere alla segmentazione interna della cavità celomatica. Questo adattamento probabilmente aiuta le sanguisughe ad allungarsi quando ingeriscono grandi quantità di sangue dai vertebrati ospiti, una condizione in cui si dice che siano "ingorgate". La sottoclasse Brachiobdella comprende minuscoli vermi simili a sanguisughe che si attaccano alle branchie o alla superficie corporea dei gamberi.
| a | b | c |
|---|---|---|
_and_Lumbricus_terrestris_(right).png) |
 |
 |
| Gruppi di anellidi. Il (a) lombrico, (b) la sanguisuga e (c) il verme piumato sono tutti anellidi. Il lombrico e la sanguisuga sono oligocheti, mentre il verme pavone è un polichete filtratore che vive in tubi. | ||
Superphylum Ecdysozoa: Nematodi e Tardigradi
modificaIl superphylum Ecdysozoa contiene un numero incredibilmente elevato di specie. Questo perché contiene due dei gruppi animali più diversi: il phylum Nematoda (gli ascaridi) e il phylum Arthropoda (gli artropodi). La caratteristica distintiva più importante degli ecdysozoi è la cuticola, un esoscheletro resistente ma flessibile che protegge questi animali dalla perdita di acqua, dai predatori e da altri pericoli dell'ambiente esterno. Un piccolo phylum all'interno degli Ecdysozoa, con un'eccezionale resistenza alla disidratazione e ad altri pericoli ambientali, è il Tardigrada. I nematodi, i tardigradi e gli artropodi appartengono tutti al superphylum Ecdysozoa, che si ritiene sia monofiletico , un clade costituito da tutti i discendenti evolutivi di un antenato comune. Tutti i membri di questo superphylum attraversano periodicamente un processo di muta che culmina nell'ecdisi, ovvero la vera e propria perdita del vecchio esoscheletro. (Il termine "ecdisi" si traduce approssimativamente come "togliere" o "spogliare"). Durante il processo di muta, la vecchia cuticola viene sostituita da una nuova cuticola, che viene secreta sotto di essa e che durerà fino al successivo periodo di crescita.
Phylum dei Nematodi
modificaI Nematoda, come altri membri del superphylum Ecdysozoa, sono triploblastici e possiedono un mesoderma embrionale che è inserito tra l'ectoderma e l'endoderma. Sono anche bilateralmente simmetrici, il che significa che una sezione longitudinale li dividerà in un lato destro e uno sinistro che sono superficialmente simmetrici. A differenza dei vermi piatti, i nematodi sono pseudocelomati e mostrano una morfologia tubulare e una sezione trasversale circolare. I nematodi includono sia forme libere che parassite.
Nel 1914, NA Cobb disse: "In breve, se tutta la materia nell'universo, eccetto i nematodi, venisse spazzata via, il nostro mondo sarebbe ancora vagamente riconoscibile e se, come spiriti disincarnati, potessimo quindi indagarlo, troveremmo le sue montagne, colline, valli, fiumi, laghi e oceani rappresentati da una sottile pellicola di nematodi..." Per parafrasare Cobb, i nematodi sono così abbondanti che se tutta la materia non nematodica della biosfera venisse rimossa, rimarrebbe ancora un'ombra del mondo precedente delineato dai nematodi! 1 Il phylum Nematoda comprende più di 28.000 specie, di cui circa 16.000 sono di natura parassitaria. Tuttavia, i nematologi ritengono che possano esserci oltre un milione di specie non classificate.
Il nome Nematoda deriva dalla parola greca "Nemos", che significa "filo", e include tutti i veri vermi tondi. I nematodi sono presenti in tutti gli habitat, in genere con ogni specie presente in grande abbondanza. Il nematode a vita libera, Caenorhabditis elegans , è stato ampiamente utilizzato come sistema modello per molti diversi filoni di indagine biologica nei laboratori di tutto il mondo.
Morfologia
modificaLa forma cilindrica del corpo dei nematodi è visibile nella Figura sotto . Questi animali hanno un sistema digerente completo con una bocca e un ano distinti, mentre nel tratto digerente dei vermi piatti è presente solo un'apertura. La bocca si apre in una faringe e un intestino muscolari, che portano a un retto e a un'apertura anale all'estremità posteriore. L'epidermide può essere un singolo strato di cellule o un sincizio , un tessuto multinucleato che in questo caso è formato dalla fusione di molte cellule singole. La cuticola dei nematodi è ricca di collagene e di un polimero chiamato chitina , che forma un'armatura protettiva all'esterno dell'epidermide. La cuticola si estende in entrambe le estremità del tratto digerente, la faringe e il retto. Nella testa, un'apertura anteriore della bocca è composta da tre (o sei) "labbra" e denti derivati dalla cuticola (in alcune specie). Alcuni nematodi possono presentare altre modifiche della cuticola come anelli, scudi della testa o verruche. Questi anelli esterni, tuttavia, non riflettono la vera segmentazione interna del corpo, che come abbiamo visto è un segno distintivo del phylum Annelida. L'attaccamento dei muscoli dei nematodi differisce da quello della maggior parte degli animali: hanno solo uno strato longitudinale e il loro attaccamento diretto ai cordoni nervosi dorsali e ventrali crea una forte contrazione muscolare che si traduce in un movimento del corpo simile a una frusta, quasi spastico.
| a |
|---|
 |
| b |
 |
 |
 |
| Morfologia dei nematodi. La micrografia elettronica a scansione mostra (a) il nematode cistico della soia ( Heterodera glycines ) e un uovo di nematode. (b) Una rappresentazione schematica mostra l'anatomia di un tipico nematode (Caenorhabditis elegans). Sotto: vista laterale (sinistro) di un ermafrodita adulto. |
Sistema escretore
modificaNei nematodi, i sistemi escretori specializzati non sono ben sviluppati. I rifiuti azotati, in gran parte sotto forma di ammoniaca , vengono rilasciati direttamente attraverso la parete corporea. In alcuni nematodi, l'osmoregolazione e l'equilibrio salino sono eseguiti da semplici cellule escretrici o ghiandole che possono essere collegate a canali accoppiati che rilasciano i rifiuti attraverso un poro anteriore. Nei nematodi marini, le cellule escretrici sono chiamate cellule renette , che sono esclusive dei nematodi.
Sistema nervoso
modificaLa maggior parte dei nematodi ha quattro cordoni nervosi longitudinali che corrono lungo la lunghezza del corpo in posizione dorsale, ventrale e laterale. Il cordone nervoso ventrale è meglio sviluppato rispetto ai cordoni dorsali e laterali. Tuttavia, tutti i cordoni nervosi si fondono all'estremità anteriore, per formare un anello nervoso faringeo attorno alla faringe, che funge da ganglio della testa o "cervello" del verme rotondo. Una fusione simile forma un ganglio posteriore sulla coda. In C. elegans , il sistema nervoso rappresenta quasi un terzo del numero totale di cellule nell'animale!
Riproduzione
modificaI nematodi impiegano una varietà di strategie riproduttive che vanno da monoiche a dioiche a partenogenetiche, a seconda della specie. C. elegans è una specie per lo più monoica con ermafroditi autofecondanti e alcuni maschi. Negli ermafroditi, ovuli e spermatozoi si sviluppano in tempi diversi nella stessa gonade. Gli ovuli sono contenuti in un utero e gli spermatozoi ameboidi sono contenuti in una spermateca ("ricettacolo dello sperma"). L'utero ha un'apertura esterna nota come vulva. Il poro genitale femminile è vicino alla parte centrale del corpo, mentre il poro genitale maschile è più vicino alla punta. Nei maschi anatomici, strutture specializzate chiamate spicole copulatorie sulla coda del maschio lo mantengono in posizione e aprono la vulva della femmina in cui lo spermatozoo ameboide viaggia nella spermateca.
La fecondazione è interna e lo sviluppo embrionale inizia molto presto dopo la fecondazione. L'embrione viene rilasciato dalla vulva durante la fase di gastrulazione. La fase di sviluppo embrionale dura 14 ore; lo sviluppo continua quindi attraverso quattro stadi larvali successivi con muta ed ecdisi che si verificano tra ogni stadio (L1, L2, L3 e L4), portando infine allo sviluppo di un giovane verme adulto. Condizioni ambientali avverse come il sovraffollamento o la mancanza di cibo possono causare la formazione di uno stadio larvale intermedio noto come larva dauer . Una caratteristica insolita di alcuni nematodi è eutely: il corpo di una data specie contiene un numero specifico di cellule come conseguenza di un rigido percorso di sviluppo.
C. elegans: il sistema modello per collegare gli studi sullo sviluppo con la genetica
modificaSe i biologi volessero studiare come si sviluppa la dipendenza dalla nicotina nel corpo, come vengono regolati i lipidi o osservare le proprietà attrattive o repellenti di certi odori, avrebbero chiaramente bisogno di progettare tre esperimenti molto diversi. Tuttavia, potrebbero aver bisogno di un solo soggetto di studio: Caenorhabditis elegans . Il nematode C. elegans è stato portato al centro della ricerca biologica tradizionale dal dott. Sydney Brenner. Dal 1963, il dott. Brenner e scienziati di tutto il mondo hanno utilizzato questo animale come sistema modello per studiare molti diversi meccanismi fisiologici e di sviluppo.
C. elegans è un nematode che vive libero nel terreno. Lungo solo circa un millimetro, può essere coltivato su piastre di agar (10.000 vermi/piastra!), nutrendosi del comune batterio intestinale Escherichia coli (un altro residente di lunga data dei laboratori biologici in tutto il mondo), e quindi può essere facilmente coltivato e mantenuto in laboratorio. Il vantaggio più grande di questo nematode è la sua trasparenza, che aiuta i ricercatori a osservare e monitorare facilmente i cambiamenti all'interno dell'animale. È anche un organismo semplice con circa 1.000 cellule e un genoma di soli 20.000 geni. I suoi cromosomi sono organizzati in cinque coppie di autosomi più una coppia di cromosomi sessuali, il che lo rende un candidato ideale per studiare la genetica. Poiché ogni cellula può essere visualizzata e identificata, questo organismo è utile per studiare fenomeni cellulari come interazioni cellula-cellula, determinazioni del destino cellulare, divisione cellulare, apoptosi (morte cellulare) e trasporto intracellulare.
Un altro enorme vantaggio è il breve ciclo di vita di questo verme ( Figura sotto). Ci vogliono solo tre giorni per raggiungere il "da uovo ad adulto a uovo figlia"; quindi, le conseguenze sullo sviluppo dei cambiamenti genetici possono essere rapidamente identificate. La durata totale della vita di C . elegans è di due o tre settimane; quindi, anche i fenomeni legati all'età sono facili da osservare. Ci sono due sessi in questa specie: ermafroditi (XX) e maschi (XO). Tuttavia, i maschi anatomici sono relativamente raramente ottenuti da accoppiamenti tra ermafroditi, poiché la loro composizione cromosomica XO richiede la non disgiunzione meiotica quando entrambi i genitori sono XX. Un'altra caratteristica che rende C . elegans un eccellente modello sperimentale è che la posizione e il numero delle 959 cellule presenti negli ermafroditi adulti di questo organismo sono costanti . Questa caratteristica è estremamente significativa quando si studia la differenziazione cellulare, la comunicazione cellula-cellula e l'apoptosi. Infine, C . elegans è anche suscettibile di manipolazioni genetiche mediante metodi molecolari, il che ne completa l'utilità come sistema modello.
I biologi di tutto il mondo hanno creato banche dati informative e gruppi dedicati alla ricerca utilizzando C. elegans . Le loro scoperte hanno portato, ad esempio, a una migliore comprensione della comunicazione cellulare durante lo sviluppo, della segnalazione neuronale e a una comprensione della regolazione dei lipidi (che è importante per affrontare problemi di salute come lo sviluppo di obesità e diabete). Negli ultimi anni, gli studi sulle ciglia in C. elegans hanno illuminato la comunità medica con una migliore comprensione della malattia renale policistica. Questo semplice organismo ha portato i biologi a scoperte complesse e significative, facendo crescere il campo della scienza in modi che toccano il mondo di tutti i giorni.

Nematodi parassiti
modificaDiversi nematodi parassiti comuni sono esempi principali di parassitismo (endoparassitismo). Questi animali importanti dal punto di vista economico e medico presentano cicli di vita complessi che spesso coinvolgono più ospiti e possono avere impatti medici e veterinari significativi. Ecco un elenco parziale di nematodi pericolosi: gli esseri umani possono essere infettati da Dracunculus medinensis , noti come vermi di Guinea, quando bevono acqua non filtrata contenente copepodi ( Figura sotto), un ospite intermedio crostaceo. Gli anchilostomi, come Ancylostoma e Necator , infestano l'intestino e si nutrono del sangue dei mammiferi, in particolare di cani, gatti ed esseri umani. I vermi della trichina ( Trichinella ) sono l'organismo causale della trichinosi negli esseri umani, spesso derivante dal consumo di carne di maiale poco cotta; la Trichinella può infettare anche altri ospiti mammiferi. L'Ascaris , un grosso verme cilindrico intestinale, ruba il nutrimento al suo ospite umano e può creare un blocco fisico dell'intestino. I vermi filariali, come Dirofilaria e Wuchereria , sono comunemente trasmessi dalle zanzare, che trasmettono gli agenti infettivi tra i mammiferi attraverso la loro attività di succhiasangue. Una specie, Wuchereria bancrofti , infetta i linfonodi di oltre 120 milioni di persone in tutto il mondo, producendo solitamente una condizione non letale ma deformante chiamata elefantiasi . In questa malattia, parti del corpo spesso si gonfiano fino a raggiungere proporzioni gigantesche a causa dell'ostruzione del drenaggio linfatico, dell'infiammazione dei tessuti linfatici e del conseguente edema. Dirofilaria immitis , un parassita infettivo del sangue, è la famigerata specie di filariosi canina.
| a |
|---|
 |
| b |
 |
| Ciclo di vita del verme di Guinea. Il verme di Guinea Dracunculus medinensis infetta circa 3,5 milioni di persone all'anno, principalmente in Africa. (a) Qui, il verme è avvolto attorno a un bastone in modo che possa essere estratto lentamente. (b) L'infezione avviene quando le persone consumano acqua contaminata da copepodi infetti, ma questo può essere facilmente prevenuto con semplici sistemi di filtraggio. |
Phylum Tardigrada
modificaI tardigradi ("slow-steppers") comprendono un phylum di piccoli animali poco appariscenti che vivono in ambienti marini, d'acqua dolce o terrestri umidi in tutto il mondo. Sono comunemente chiamati "orsi d'acqua" per via dei loro corpi paffuti e dei grandi artigli sulle loro zampe tozze. Esistono oltre 1.000 specie, la maggior parte delle quali è lunga meno di 1 mm. Una cuticola chitinosa ricopre la superficie del corpo e può essere divisa in placche ( Figura sotto). I tardigradi sono noti per la loro capacità di entrare in uno stato chiamato criptobiosi , che fornisce loro resistenza a molteplici sfide ambientali, tra cui disidratazione, temperature molto basse, vuoto, alta pressione e radiazioni. Possono sospendere la loro attività metabolica per anni e sopravvivere alla perdita fino al 99% del loro contenuto d'acqua. La loro notevole resistenza è stata recentemente attribuita a proteine uniche che sostituiscono l'acqua nelle loro cellule e proteggono la loro struttura cellulare interna e il loro DNA dai danni.

Morfologia e Fisiologia
modificaI tardigradi hanno corpi cilindrici, con quattro paia di zampe che terminano con un certo numero di artigli. La cuticola viene periodicamente persa, compresa la cuticola che ricopre gli artigli. Le prime tre paia di zampe sono usate per camminare e la coppia posteriore per aggrapparsi al substrato. Una bocca circolare conduce a una faringe muscolare e a ghiandole salivari. I tardigradi si nutrono di piante, alghe o piccoli animali. Le cellule vegetali vengono perforate con uno stiletto chitinoso e il contenuto cellulare viene poi risucchiato nell'intestino dalla faringe muscolare. Fasce di singole cellule muscolari sono attaccate ai vari punti dell'epidermide e si estendono nelle zampe per fornire movimento ambulatoriale. La cavità corporea principale è un emocele , ma non ci sono strutture circolatorie specializzate per muovere il sangue, né ci sono strutture respiratorie specializzate. I tubuli malpighiani nell'emocele rimuovono i rifiuti metabolici e li trasportano nell'intestino. Un cervello dorsale è collegato a un cordone nervoso ventrale con gangli segmentali associati alle appendici. Le strutture sensoriali sono notevolmente ridotte, ma c'è un paio di semplici macchie oculari sulla testa e ciglia sensoriali o setole concentrate verso l'estremità della testa dell'animale.
Riproduzione
modificaLa maggior parte dei tardigradi sono dioici e sia i maschi che le femmine hanno una sola gonade. L'accoppiamento avviene solitamente al momento della muta e la fecondazione è esterna. Le uova possono essere deposte in una cuticola muta o attaccate ad altri oggetti. Lo sviluppo è diretto e l'animale può mutare una dozzina di volte durante la sua vita. Nei tardigradi, come i nematodi, lo sviluppo produce un numero fisso di cellule, con il numero effettivo di cellule che dipende dalla specie. Un'ulteriore crescita avviene ingrandendo le cellule, non moltiplicandole.
Superphylum Ecdysozoa: Artropodi
modificaIl superphylum Ecdysozoa include anche il phylum Arthropoda, uno dei cladi di animali di maggior successo sul pianeta. Gli artropodi sono organismi celomati caratterizzati da un robusto esoscheletro chitinoso e appendici articolate. Sono state descritte ben oltre un milione di specie di artropodi e i sistematici ritengono che ci siano milioni di specie in attesa di una classificazione adeguata. Come altri Ecdysozoa, tutti gli artropodi attraversano periodicamente il processo fisiologico della muta , seguito dall'ecdisi (la vera e propria perdita dell'esoscheletro), mentre crescono. Gli artropodi sono organismi eucoelomati, protostomici, spesso con cicli di vita incredibilmente complicati.
Phylum degli Artropodi
modificaIl nome "arthropoda" significa "piedi articolati". Il nome descrive appropriatamente gli invertebrati inclusi in questo phylum. Gli artropodi hanno probabilmente sempre dominato il regno animale in termini di numero di specie e probabilmente continueranno a farlo: si stima che l'85 percento di tutte le specie conosciute siano incluse in questo phylum! In effetti, la vita sulla Terra potrebbe plausibilmente essere definita l'era degli artropodi iniziata circa 500 milioni di anni fa.
Le caratteristiche principali di tutti gli animali di questo phylum sono la segmentazione strutturale e funzionale del corpo e la presenza di appendici articolate. Gli artropodi hanno un esoscheletro costituito principalmente da chitina, un polisaccaride impermeabile e resistente composto da N-acetilglucosamina. Il phylum Arthropoda è il clade più specifico del mondo animale ( Tabella 28.1 ), e gli insetti formano la classe più grande all'interno di questo phylum. Per un confronto, fare riferimento al numero approssimativo di specie nei phyla elencati di seguito.
| Tipologia | # specie |
| Ctenofori | 100 |
| Poriferi | 5.000 |
| Cnidari | 11.000 |
| Platelminti | 25.000 |
| Rotifera | 2.000 |
| Nemertea | 1.200 |
| Anellidi | 22.000 |
| Mollusco | 112.000 |
| Nematodi | 28.000+ |
| Tardigradi | >1.000 |
| Artropodi | 1.134.000 |
| Echinodermi | 7.000 |
| Cordati | 100.000 |
Tavolo 28.1
Il phylum Arthropoda comprende animali che hanno avuto successo nella colonizzazione di habitat terrestri, acquatici e aerei. Questo phylum è ulteriormente classificato in cinque subphyla: Trilobita (trilobiti, tutti estinti), Chelicerata (limuli, ragni, scorpioni, zecche, acari e daddy longlegs o harvestmen), Myriapoda (millepiedi, centopiedi e loro parenti), Crustacea (granchi, aragoste, gamberi, isopodi, cirripedi e un po' di zooplancton) ed Hexapoda (insetti e loro parenti a sei zampe). I trilobiti, un gruppo estinto di artropodi rinvenuto principalmente durante l'era Cambriana (circa 500 milioni di anni fa), sono probabilmente i più strettamente imparentati con i Chelicerata. Questi sono identificati in base ai loro fossili; erano piuttosto diversificati e si irradiarono in modo significativo in migliaia di specie prima della loro completa estinzione alla fine del Permiano, circa 240 milioni di anni fa ( Figura sotto).

Morfologia
modificaLe caratteristiche caratteristiche degli artropodi includono la presenza di appendici articolate, segmentazione del corpo ed esoscheletro chitinizzato. La fusione di gruppi adiacenti di segmenti ha dato origine a regioni funzionali del corpo chiamate tagmata (singolare = tagma). I tagmata possono avere la forma di testa, torace e addome, o cefalotorace e addome, o testa e tronco, a seconda del taxon. I tagmata comunemente descritti possono essere composti da un numero diverso di segmenti; ad esempio, la testa della maggior parte degli insetti risulta dalla fusione di sei segmenti ancestrali, mentre la "testa" di un altro artropode può essere composta da meno segmenti ancestrali, a causa di eventi evolutivi indipendenti. Le appendici articolate degli artropodi, spesso in coppie segmentali, sono state specializzate per varie funzioni: percepire il loro ambiente (antenne), catturare e manipolare il cibo (mandibole e mascelle), nonché camminare, saltare, scavare e nuotare.
Nel corpo dell'artropode è presente una cavità centrale, chiamata emocele (o cavità sanguigna), e i fluidi dell'emocele vengono mossi dalla contrazione di regioni del vaso sanguigno dorsale tubulare chiamate "cuori". I gruppi di artropodi differiscono anche negli organi utilizzati per l'escrezione di rifiuti azotati, con i crostacei che possiedono ghiandole verdi e gli insetti che utilizzano tubuli malpighiani , che lavorano insieme all'intestino posteriore per riassorbire l'acqua mentre liberano il corpo dai rifiuti azotati. Il sistema nervoso tende a essere distribuito tra i segmenti, con gangli più grandi in segmenti con strutture sensoriali o appendici. I gangli sono collegati da un cordone nervoso ventrale.
I sistemi respiratori variano a seconda del gruppo di artropodi. Insetti e miriapodi utilizzano una serie di tubi (trachee) che si diramano attraverso il corpo, terminando in minuscole tracheole. Questi sottili tubi respiratori eseguono lo scambio di gas direttamente tra l'aria e le cellule all'interno dell'organismo. Le trachee principali si aprono sulla superficie della cuticola tramite aperture chiamate spiracoli. Dovremmo notare che questi sistemi tracheali di ventilazione si sono evoluti indipendentemente negli esapodi, nei miriapodi e negli aracnidi. Sebbene il sistema tracheale funzioni estremamente bene negli ambienti terrestri, funziona bene anche negli ambienti acquatici di acqua dolce: infatti, numerose specie di insetti acquatici sia negli stadi immaturi che in quelli adulti possiedono sistemi tracheali. Tuttavia, sebbene vi siano insetti che vivono sulla superficie degli ambienti marini, nessuno è strettamente marino, il che significa che completano la loro intera metamorfosi in acqua salata.
Al contrario, i crostacei acquatici utilizzano le branchie, i chelicerati terrestri utilizzano polmoni a libro e i chelicerati acquatici utilizzano branchie a libro ( Figura sotto). I polmoni a libro degli aracnidi (scorpioni, ragni, zecche e acari) contengono una pila verticale di tessuto della parete dell'emocele che assomiglia in qualche modo alle pagine di un libro. Tra ciascuna delle "pagine" di tessuto c'è uno spazio d'aria. Ciò consente a entrambi i lati del tessuto di essere sempre a contatto con l'aria, aumentando notevolmente l'efficienza dello scambio di gas. Le branchie dei crostacei sono strutture filamentose che scambiano gas con l'acqua circostante.
La cuticola è la "copertura" dura di un artropode. È composta da due strati: l' epicuticola , che è uno strato esterno sottile, ceroso e resistente all'acqua che non contiene chitina, e lo strato sottostante, la procuticola chitinosa , che a sua volta è composta da un'esocuticola e da un'endocuticola inferiore , tutte supportate in ultima analisi da una membrana basale . L'esoscheletro è molto protettivo (a volte è difficile schiacciare un grosso coleottero!), ma non sacrifica flessibilità o mobilità. Sia l'esocuticola (che viene secreta prima di una muta), sia un'endocuticola (che viene secreta dopo una muta), sono composte da chitina legata a una proteina; la chitina è insolubile in acqua, alcali e acidi deboli. La procuticola non è solo flessibile e leggera, ma fornisce anche protezione contro la disidratazione e altri stress biologici e fisici. Alcuni artropodi, come i crostacei, aggiungono sali di calcio al loro esoscheletro, il che aumenta la resistenza della cuticola, ma ne riduce la flessibilità. In alcuni casi, come nelle aragoste, la quantità di sale di calcio depositato nella chitina è estrema. Per crescere, l'artropode deve "sbarazzarsi" dell'esoscheletro durante il processo fisiologico chiamato muta , seguito dall'effettiva rimozione della cuticola esterna, chiamata ecdisi ("spogliarsi"). All'inizio, questo sembra essere un metodo di crescita pericoloso, perché mentre il nuovo esoscheletro si indurisce, l'animale è vulnerabile alla predazione; tuttavia, la muta e l'ecdisi consentono anche la crescita e il cambiamento della morfologia, nonché una grande diversificazione delle dimensioni, semplicemente perché il numero di mute può essere modificato attraverso l'evoluzione.
Di seguito viene descritta la morfologia caratteristica degli animali rappresentativi di ciascun subphylum.
| a |
|---|
 |
 |
| b |
 |
| Strutture respiratorie degli artropodi. I polmoni a libro degli aracnidi (a) sono costituiti da sacche d'aria alternate e tessuto emocele a forma di pila di libri (da cui il nome, "polmone a libro"). Le branchie a libro dei chelicerati: (b), nei limuli sono simili ai polmoni a libro ma sono esterne in modo che lo scambio di gas possa avvenire con l'acqua circostante (nella foto quelle al centro più chiare) |
Subphylum Chelicerati
modificaQuesto subphylum include animali come granchi ferro di cavallo, ragni marini, ragni, acari, zecche, scorpioni, scorpioni frusta e opilioni. I chelicerati sono prevalentemente terrestri, sebbene esistano anche alcune specie di acqua dolce e marine. Si stima che 77.000 specie di chelicerati possano essere trovate in quasi tutti gli habitat terrestri.
Il corpo dei chelicerati è diviso in due tagmi: prosoma e opistosoma, che sono fondamentalmente gli equivalenti di un cefalotorace (solitamente più piccolo) e di un addome (solitamente più grande). Un tagma "testa" distinto non è solitamente discernibile. Il phylum deriva il suo nome dalla prima coppia di appendici: i cheliceri ( Figura sotto), che servono come apparati boccali specializzati simili ad artigli o zanne. Dovremmo notare qui che i cheliceri sono in realtà zampe modificate , ma non sono l'esatto equivalente seriale delle mandibole, che sono gli apparati boccali masticatori simili a zampe modificati di insetti e crostacei: i cheliceri sono portati sul primo segmento che costituisce il prosoma, mentre le mandibole sono embrionicamente sul quarto segmento della testa mandibolare. I cheliceri hanno perso secondariamente le loro antenne e quindi non le hanno.

Alcune delle funzioni delle antenne (come il tatto) sono ora svolte dal secondo paio di appendici, i pedipalpi , che possono anche essere utilizzati per la percezione generale dell'ambiente e per la manipolazione del cibo. In alcune specie, come i ragni marini, è presente un ulteriore paio di appendici derivate delle zampe, chiamate ovigeri, tra i cheliceri e i pedipalpi. Gli ovigeri sono utilizzati per la pulizia e dai maschi per trasportare le uova. Nei ragni, i cheliceri sono spesso modificati e terminano in zanne che iniettano veleno nelle loro prede prima di nutrirsi ( Figura sotto).
La maggior parte dei chelicerati ingerisce il cibo utilizzando una cavità preorale formata dai cheliceri e dai pedipalpi. Alcuni chelicerati possono secernere enzimi digestivi per predigerire il cibo prima di ingerirlo. I chelicerati parassiti come zecche e acari hanno sviluppato apparati succhiasangue. I membri di questo subphylum hanno un sistema circolatorio aperto con un cuore che pompa il sangue nell'emocele. Le specie acquatiche, come i limuli, hanno branchie, mentre le specie terrestri hanno trachee o polmoni a libro per lo scambio gassoso. L'emolinfa dei chelicerati contiene emocianina, una proteina di trasporto dell'ossigeno contenente rame.
Il sistema nervoso nei chelicerati è costituito da un cervello e due cordoni nervosi ventrali. I chelicerati sono dioici, ovvero i sessi sono separati. Questi animali utilizzano strategie di fecondazione esterna e interna per la riproduzione, a seconda della specie e del suo habitat. Le cure parentali per i piccoli variano da assolutamente nessuna a cure relativamente prolungate.
Collegamento all'apprendimento
modificaVisita questo sito per leggere una lezione sugli artropodi, tra cui mappe interattive dell'habitat e molto altro.
Subphylum Miriapodi
modificaIl subphylum Myriapoda comprende artropodi con numerose zampe. Sebbene il nome sia fuorviante, suggerendo che in questi invertebrati siano presenti migliaia di zampe, il numero di zampe varia in genere da 10 a 750. Questo subphylum comprende 16.000 specie; gli esempi più comuni sono i millepiedi e i centopiedi. Praticamente tutti i miriapodi sono animali terrestri e preferiscono un ambiente umido. Gli antichi miriapodi (o artropodi simili ai miriapodi) dal Siluriano al Devoniano crescevano fino a 10 piedi di lunghezza (tre metri). Sfortunatamente, sono tutti estinti!
I miriapodi si trovano tipicamente in terreni umidi, materiale biologico in decomposizione e lettiera di foglie. Il subphylum Myriapoda è diviso in quattro classi: Chilopoda, Symphyla, Diplopoda e Pauropoda. I centopiedi come Scutigera coleoptrata ( Figura sotto a) sono classificati come chilopodi. Questi animali hanno un paio di zampe per segmento, mandibole come apparato boccale e sono in qualche modo appiattiti dorsoventralmente. Tutti i centopiedi sono predatori e le zampe nel primo segmento sono modificate per formare forcipule (artigli velenosi) che rilasciano veleno alle prede. I Symphyla sono simili ai centopiedi, ma non hanno gli artigli velenosi e sono vegetariani. I millepiedi hanno due paia di zampe per diplosegmento, una caratteristica che deriva dalla fusione embrionale di coppie adiacenti di segmenti corporei. Questi artropodi hanno solitamente una sezione trasversale più rotonda rispetto ai millepiedi e sono erbivori o detritivori. I millepiedi hanno due paia di zampe per segmento, posizionate sotto il corpo, rispetto ai millepiedi, che ne hanno una coppia per segmento, posizionata lateralmente al corpo ( Figura sotto b). I millepiedi si nutrono di materiale vegetale in decomposizione, mentre i millepiedi sono predatori e mangiano insetti che uccidono con il loro veleno. I Pauropodi sono simili ai millepiedi, ma hanno meno segmenti.
| a | b |
|---|---|
 |
 |
| Miriapodi. Il centopiedi Scutigera coleoptrata (a) ha fino a 15 paia di zampe. Il millepiedi nordamericano Narceus americanus (b) ha molte zampe, anche se non mille, come potrebbe suggerire il suo nome. | |
Subphylum Crostacei
modificaI crostacei sono gli artropodi acquatici (sia d'acqua dolce che marina) più dominanti, con un numero totale di crostacei marini di circa 70.000 specie. Krill, gamberi, aragoste, granchi e gamberi di fiume sono esempi di crostacei ( Figura sotto). Tuttavia, ci sono anche numerose specie di crostacei terrestri: specie terrestri come i pidocchi del legno ( Armadillidium spp.), chiamati anche porcellini di terra, roly-polies, insetti della patata o isopodi, sono anch'essi crostacei. Tuttavia, il numero di specie terrestri in questo subphylum è relativamente basso.
| a | b | c |
|---|---|---|
 |
 |
 |
| Crostacei. Il (a) granchio e (b) gambero krill sono entrambi crostacei acquatici. Il porcellino di terra Armadillidium è un crostaceo terrestre | ||
I crostacei possiedono tipicamente due paia di antenne, mandibole come apparato boccale e appendici biramose ("biramificate"), il che significa che le loro zampe sono formate in due parti chiamate endopodi ed esopodi, che sembrano superficialmente distinte dalle zampe uniramiche ("un ramo") dei miriapodi e degli esapodi ( Figura sotto). Poiché le appendici biramose sono presenti anche nei trilobiti, le appendici biramose rappresentano la condizione ancestrale negli artropodi. Attualmente, descriviamo vari artropodi come aventi appendici uniramiche o biramose, ma queste sono solo descrittive e non riflettono necessariamente relazioni evolutive diverse dal fatto che tutte le zampe articolate degli artropodi condividono un'ascendenza comune.

Nella maggior parte dei crostacei, la testa e il torace sono fusi per formare un cefalotorace ( Figura sotto), che è coperto da una piastra chiamata carapace , producendo così un piano corporeo comprendente due tagmata: cefalotorace e addome . I crostacei hanno un esoscheletro chitinoso che viene perso tramite muta ed ecdisi ogni volta che l'animale richiede un aumento di dimensioni o la fase successiva di sviluppo. Gli esoscheletri di molte specie acquatiche sono anche infusi di carbonato di calcio, che li rende ancora più forti di quelli di altri artropodi. I crostacei hanno un sistema circolatorio aperto in cui il sangue viene pompato nell'emocele dal cuore situato dorsalmente. L'emocianina è il principale pigmento respiratorio presente nei crostacei, ma l'emoglobina si trova in alcune specie ed entrambi sono disciolti nell'emolinfa anziché trasportati nelle cellule.

Come nei chelicerati, la maggior parte dei crostacei è dioica. Tuttavia, alcune specie come i cirripedi possono essere ermafroditi. L'ermafroditismo seriale , in cui la gonade può passare dalla produzione di sperma a quella di ovuli, è anch'esso esibito in alcune specie. Le uova fecondate possono essere trattenute all'interno della femmina della specie o possono essere rilasciate nell'acqua. I crostacei terrestri cercano spazi umidi nei loro habitat per deporre le uova.
Gli stadi larvali, nauplius o zoea, sono visibili nello sviluppo iniziale dei crostacei acquatici. Una larva di cypris è visibile anche nello sviluppo iniziale dei cirripedi ( Figura sotto ).
| a | b | c |
|---|---|---|
 |
 |
 |
| Larve di crostacei. Tutti i crostacei attraversano diversi stadi larvali. Sono mostrati (a) lo stadio larvale nauplius di un gambero girino, (b) lo stadio larvale cypris di un cirripede e (c) lo stadio larvale zoea di un granchio verde | ||
I crostacei possiedono un cervello formato dalla fusione dei primi tre gangli segmentali, così come due occhi composti. Un cordone nervoso ventrale collega gangli segmentali aggiuntivi. La maggior parte dei crostacei è carnivora, ma sono note specie erbivore e detritivore, e persino specie endoparassite. Una specie endoparassite altamente evoluta, come Sacculina spp., parassita il suo ospite granchio e alla fine lo distrugge dopo averlo costretto a covare le uova del parassita! I crostacei possono anche essere cannibali quando sono presenti popolazioni estremamente elevate di questi organismi.
Subphylum Esapodi
modificaGli insetti costituiscono la classe più numerosa di artropodi in termini di diversità di specie e di biomassa, almeno negli habitat terrestri.
Il nome Hexapoda descrive la presenza di sei zampe (tre paia) in questi animali, che li differenzia da altri gruppi di artropodi che hanno un numero diverso di zampe. In alcuni casi, tuttavia, il numero di zampe è stato ridotto evolutivamente, oppure le zampe sono state altamente modificate per adattarsi a condizioni specifiche, come l'endoparassitismo. I corpi degli esapodi sono organizzati in tre tagmata: testa, torace e addome. I singoli segmenti della testa hanno parti della bocca derivate da zampe articolate, e il torace ha tre paia di appendici articolate, e anche ali, nella maggior parte dei gruppi derivati. Ad esempio, negli pterigoti (insetti alati), oltre a una coppia di zampe articolate su tutti e tre i segmenti che compongono il torace (protorace, mesotorace e metatorace), ci sono ali venate sul mesotorace e sul metatorace.
Anche le appendici presenti su altri segmenti corporei derivano evolutivamente da zampe modificate. In genere, la testa presenta un "labbro" superiore o labbro e mandibole (o derivazione delle mandibole) che fungono da apparato boccale; mascelle e un "labbro" inferiore chiamato labbro: entrambi manipolano il cibo. La testa presenta anche un paio di antenne sensoriali, nonché organi sensoriali quali un paio di occhi composti, ocelli (occhi semplici) e numerosi peli sensoriali. L'addome presenta solitamente 11 segmenti e presenta aperture riproduttive esterne. Il subphylum Hexapoda comprende alcuni insetti alati (come i moscerini della frutta) e altri secondariamente privi di ali (come le pulci). L'unico ordine di insetti "primitivamente privi di ali" è il Thysanura, i Bristletail. Tutti gli altri ordini sono alati o discendenti di insetti formalmente alati.
L'evoluzione delle ali è un mistero importante e irrisolto. A differenza dei vertebrati, le cui "ali" sono semplicemente preadattamenti di "braccia" che servivano come fondamenta strutturali per l'evoluzione delle ali funzionali (ciò è avvenuto indipendentemente negli pterosauri, nei dinosauri [uccelli] e nei pipistrelli), l'evoluzione delle ali negli insetti è ciò che chiamiamo uno sviluppo de novo (nuovo) che ha dato agli pteriogoti il dominio sulla Terra. Gli insetti alati esistevano oltre 425 milioni di anni fa e, nel Carbonifero, si erano evoluti diversi ordini di insetti alati ( Paleoptera ), la maggior parte dei quali ora estinti. Ci sono buone prove fisiche che le ninfe paleozoiche con alette toraciche (forse incernierate, ex opercoli branchiali di specie semi-acquatiche) usassero questi dispositivi sulla terraferma per aumentare la temperatura toracica (il torace è dove si trovano le zampe) a livelli che avrebbero consentito loro di sfuggire ai predatori più velocemente, trovare più risorse alimentari e compagni e disperdersi più facilmente. Le alette toraciche (che possono essere trovate su insetti fossilizzati precedenti all'avvento di insetti veramente alati) avrebbero potuto essere facilmente selezionate per scopi termoregolatori prima di raggiungere una dimensione che avrebbe consentito loro la capacità di planare o di volare effettivamente sbattendo le ali. Anche gli insetti moderni con ali ampiamente attaccate, come le farfalle, usano il terzo basale delle loro ali (l'area accanto al torace) per la termoregolazione e i due terzi esterni per il volo, il mimetismo e la selezione del compagno.
Molti degli insetti comuni che incontriamo quotidianamente, tra cui formiche, coleotteri, blatte, farfalle, grilli e mosche, sono esempi di Hexapoda. Tra questi, formiche, coleotteri, mosche e farfalle adulte si sviluppano tramite metamorfosi completa da larve simili a larve o bruchi, mentre blatte e grilli adulti si sviluppano tramite una metamorfosi graduale o incompleta da immaturi senza ali. Tutta la crescita avviene durante le fasi giovanili. Gli adulti non crescono ulteriormente (ma possono diventare più grandi) dopo la loro muta finale. Le variazioni nella morfologia delle ali, delle zampe e dell'apparato boccale contribuiscono tutte all'enorme varietà osservata negli insetti. La variabilità degli insetti è stata anche incoraggiata dalla loro attività di impollinatori e dalla loro coevoluzione con le piante da fiore. Alcuni insetti, in particolare termiti, formiche, api e vespe, sono eusociali, il che significa che vivono in grandi gruppi con individui assegnati a ruoli o caste specifici, come regina, fuco e operaia. Gli insetti sociali utilizzano i feromoni , segnali chimici esterni, per comunicare e mantenere la struttura del gruppo e la coesione della colonia.

Figura 28.45 Anatomia degli insetti. In questa anatomia di base di un insetto esapode, si noti che gli insetti hanno un apparato digerente ben sviluppato (verde), un apparato respiratorio (n.8 in grigio), un apparato circolatorio (rosso) e un sistema nervoso (n.5-verdeazzurro e n.28-blu). Si noti il "cuore" multiplo (n.14) e i gangli segmentali (n.28).
Esercizio: quale delle seguenti affermazioni sugli insetti è falsa?
- Gli insetti hanno vasi sanguigni sia dorsali che ventrali.
- Gli insetti sono dotati di spiracoli, aperture che consentono all'aria di entrare nel sistema tracheale.
- La trachea fa parte dell'apparato digerente.
- La maggior parte degli insetti ha un apparato digerente ben sviluppato, dotato di bocca, gozzo e intestino.
Superphylum Deuterostomia
modificaI phyla Echinodermata e Chordata (il phylum che include gli esseri umani) appartengono entrambi al superphylum Deuterostomia. Ricorda che protostomi e deuterostomi differiscono in certi aspetti del loro sviluppo embrionale e prendono il nome in base a quale apertura dell'archenteron ( tubo intestinale primitivo) si sviluppa per prima. La parola deuterostome deriva dalla parola greca che significa "bocca seconda", indicando che la bocca si sviluppa come una struttura secondaria opposta alla posizione del blastoporo, che diventa l'ano. Nei protostomi ("bocca prima"), la prima apertura embrionale diventa la bocca e la seconda apertura diventa l'ano.
Ci sono una serie di altre caratteristiche di sviluppo che differiscono tra protostomi e deuterostomi, tra cui il tipo di scissione precoce (divisione cellulare embrionale) e la modalità di formazione del celoma dell'embrione: i protosomi presentano in genere una scissione a mosaico a spirale, mentre i deuterostomi presentano una scissione regolativa radiale. Nei deuterostomi, il rivestimento endodermico dell'archenteron di solito forma gemme chiamate tasche celomiche che si espandono e alla fine cancellano il blastocele embrionale (la cavità all'interno della blastula e della gastrula precoce) per diventare il mesoderma embrionale , il terzo foglietto germinale. Ciò accade quando le tasche mesodermiche si separano dallo strato endodermico invaginante che forma l'archenteron, quindi si espandono e si fondono per formare la cavità celomica. Il celoma risultante è definito enterocele . L'archenteron si sviluppa nel canale alimentare e un'apertura della bocca è formata dall'invaginazione dell'ectoderma al polo opposto al blastoporo della gastrula. Il blastoporo forma l'ano del sistema alimentare nelle forme giovanili e adulte. La scissione nella maggior parte dei deuterostomi è anche indeterminata , il che significa che i destini evolutivi delle cellule embrionali precoci non sono decisi in quel punto dello sviluppo embrionale (questo è il motivo per cui potremmo potenzialmente clonare la maggior parte dei deuterostomi, compresi noi stessi).
I deuterostomi sono costituiti da due cladi principali: i Chordata e gli Ambulacraria. I Chordata includono i vertebrati e due subphyla di invertebrati, gli urocordati e i cefalocordati. Gli Ambulacraria includono gli echinodermi e gli emicordati, che un tempo erano considerati un subphylum di cordati ( Figura 28.46 ). I due cladi, oltre a essere deuterostomi, hanno altre interessanti caratteristiche in comune. Come abbiamo visto, la stragrande maggioranza degli animali invertebrati non possiede un endoscheletro vertebrale osseo definito, o un cranio osseo. Tuttavia, uno dei gruppi più ancestrali di invertebrati deuterostomi, gli Echinodermata, produce minuscole "ossa" scheletriche chiamate ossicini che costituiscono un vero e proprio endoscheletro , o scheletro interno, ricoperto da un'epidermide. Gli Hemichordata (vermi ghianda e pterobranchi) non saranno trattati qui, ma condividono con gli echinodermi un celoma in tre parti (tripartito), forme larvali simili e un metanefridio derivato che libera gli animali dai rifiuti azotati. Condividono anche le fessure faringee con i cordati ( Figura 28.46 ). Inoltre, gli hemichordata hanno un cordone nervoso dorsale nella linea mediana dell'epidermide, ma non hanno un tubo neurale, una vera notocorda e l'endostilo e la coda post-anale caratteristici dei cordati.

Phylum Echinodermata
modificaGli Echinodermata prendono il nome dalla loro "pelle spinosa" (dal greco "echinos" che significa "spinoso" e "dermos" che significa "pelle"). Questo phylum è una raccolta di circa 7.000 specie viventi descritte di organismi esclusivamente marini, che vivono sui fondali. Le stelle marine ( Figura sotto), i cetrioli di mare, i ricci di mare, i dollari di sabbia e le stelle fragili sono tutti esempi di echinodermi.
Morfologia e anatomia
modificaNonostante il valore adattativo della bilateralità per la maggior parte degli animali cefalizzati che vivono liberi, gli echinodermi adulti mostrano una simmetria pentaradiale (con "braccia" solitamente disposte in multipli di cinque attorno a un asse centrale). Gli echinodermi hanno un endoscheletro fatto di ossicini calcarei (piccole placche ossee), ricoperti dall'epidermide. Per questo motivo, è un endoscheletro come il nostro, non un esoscheletro come quello degli artropodi. Gli ossicini possono essere fusi insieme, incorporati separatamente nel tessuto connettivo del derma o essere ridotti a minuscole spicole ossee come nei cetrioli di mare. Le spine da cui prendono il nome gli echinodermi sono collegate ad alcune delle placche. Le spine possono essere mosse da piccoli muscoli, ma possono anche essere bloccate in posizione per difesa. In alcune specie, le spine sono circondate da minuscoli artigli peduncolati chiamati pedicellari , che aiutano a mantenere la superficie dell'animale pulita da detriti, a proteggere le papule utilizzate nella respirazione e talvolta facilitano la cattura del cibo.
L'endoscheletro è prodotto da cellule dermiche, che producono anche diversi tipi di pigmenti, conferendo colori vividi a questi animali. Nelle stelle marine, proiezioni simili a dita (papille) di tessuto dermico si estendono attraverso l'endoscheletro e funzionano come branchie. Alcune cellule sono ghiandolari e possono produrre tossine. Ogni braccio o sezione dell'animale contiene diverse strutture: ad esempio, ghiandole digestive, gonadi e i piedi tubolari che sono unici per gli echinodermi. Negli echinodermi come le stelle marine, ogni braccio porta due file di piedi tubolari sul lato orale, che corrono lungo un solco ambulacrale esterno. Questi piedi tubolari aiutano nella locomozione, nell'alimentazione e nelle sensazioni chimiche, oltre a servire per attaccare alcune specie al substrato.

Sistemi vascolari e ematici e idrologici
modificaGli echinodermi hanno un sistema ambulacrale (vascolare dell'acqua) unico , derivato da una parte del celoma , o "cavità corporea". Il sistema vascolare dell'acqua è costituito da un canale ad anello centrale e canali radiali che si estendono lungo ogni braccio. Ogni canale radiale è collegato a una doppia fila di piedi tubolari , che sporgono attraverso fori nell'endoscheletro e funzionano come strutture tattili e ambulatoriali. Questi piedi tubolari possono estendersi o ritrarsi in base al volume di acqua presente nel sistema di quel braccio, consentendo all'animale di muoversi e anche di catturare la preda con la loro azione a ventosa. I singoli piedi tubolari sono controllati da ampolle a forma di bulbo . L'acqua di mare entra nel sistema attraverso una madreporite aborale (opposta all'area orale in cui si trova la bocca) e passa al canale ad anello attraverso un breve canale di pietra . L'acqua che circola attraverso queste strutture facilita lo scambio gassoso e fornisce una fonte idrostatica per la locomozione e la manipolazione delle prede. Un sistema emale , costituito da anelli orali, gastrici e aborali, nonché altri vasi che corrono più o meno parallelamente al sistema vascolare dell'acqua, fa circolare i nutrienti. Il trasporto di nutrienti e gas è condiviso dai sistemi vascolare dell'acqua ed emale oltre alla cavità corporea viscerale che circonda gli organi principali.
Sistema nervoso
modificaIl sistema nervoso di questi animali è relativamente semplice, comprendente un anello nervoso circumorale al centro e cinque nervi radiali che si estendono verso l'esterno lungo le braccia. Inoltre, diverse reti di nervi sono situate in diverse parti del corpo. Tuttavia, in questi animali non sono presenti strutture analoghe a un cervello o a grandi gangli. A seconda del gruppo, gli echinodermi possono avere organi sensoriali ben sviluppati per il tatto e la chemiorecezione (ad esempio, all'interno dei piedi tubolari e sui tentacoli sulle punte delle braccia), così come fotorecettori e statocisti.
Apparato digerente ed escretore
modificaUna bocca, situata sul lato orale (ventrale), si apre attraverso un breve esofago verso un grande stomaco a forma di sacco. Il cosiddetto stomaco "cardiaco" può essere rovesciato attraverso la bocca durante l'alimentazione (ad esempio, quando una stella marina rovescia il suo stomaco in una preda bivalve per digerire l'animale, vivo , all'interno del suo stesso guscio!) Ci sono masse di ghiandole digestive ( ciechi pilorici ) in ogni braccio, che corrono dorsalmente lungo le braccia e ricoprono le ghiandole riproduttive sottostanti. Dopo essere passato attraverso i ciechi pilorici in ogni braccio, il cibo digerito viene incanalato verso un piccolo ano, se ne esiste uno.
I podociti, cellule specializzate per l'ultrafiltrazione dei fluidi corporei, sono presenti vicino al centro del disco dell'echinoderma, alla giunzione dei sistemi vascolare e emale dell'acqua. Questi podociti sono collegati tramite un sistema interno di canali alla madreporite , dove l'acqua entra nel canale della pietra. L'echinoderma adulto ha in genere un celoma spazioso e pieno di fluido. Le ciglia aiutano a far circolare il fluido all'interno della cavità corporea e conducono alle papule piene di fluido , dove avviene lo scambio di ossigeno e anidride carbonica, nonché la secrezione di rifiuti azotati come l'ammoniaca, per diffusione.
Riproduzione
modificaGli echinodermi sono dioici, ma maschi e femmine sono indistinguibili a parte i loro gameti. Maschi e femmine rilasciano i loro gameti nell'acqua contemporaneamente e la fecondazione è esterna. I primi stadi larvali di tutti gli echinodermi (ad esempio, la bipinnaria degli echinodermi asteroidi come le stelle marine) hanno simmetria bilaterale, sebbene ogni classe di echinodermi abbia la sua forma larvale. Le forme adulte radialmente simmetriche si formano da un gruppo di cellule nella larva. Le stelle marine, le stelle fragili e i cetrioli di mare possono anche riprodursi asessualmente per frammentazione , così come rigenerare parti del corpo perse a causa di traumi, anche quando oltre il 75 percento della loro massa corporea è persa!
Classi di Echinodermi
modificaQuesto phylum è suddiviso in cinque classi attuali: Asteroidea (stelle marine), Ophiuroidea (stelle fragili), Echinoidea (ricci di mare e dollari di sabbia), Crinoidea (gigli di mare o stelle piumate) e Holothuroidea (cetrioli di mare) ( Figura sotto).
Gli echinodermi più noti appartengono alla classe Asteroidea, o stelle marine. Hanno una grande varietà di forme, colori e dimensioni, con oltre 1.800 specie finora note. La caratteristica principale delle stelle marine che le distingue dalle altre classi di echinodermi include spesse braccia che si estendono da un disco centrale da cui vari organi del corpo si diramano nelle braccia. All'estremità di ogni braccio ci sono semplici macchie oculari e tentacoli che fungono da recettori tattili. Le stelle marine usano le loro file di piedi tubolari non solo per afferrare le superfici, ma anche per afferrare le prede. La maggior parte delle stelle marine sono carnivore e le loro prede principali appartengono al phylum Mollusca. Manipolando i suoi piedi tubolari, una stella marina può aprire i gusci dei molluschi. Le stelle marine hanno due stomaci, uno dei quali può sporgere dalla bocca e secernere succhi digestivi nella o sulla preda, anche prima dell'ingestione. Una stella marina che mangia una vongola può aprire parzialmente il guscio e quindi rovesciare lo stomaco nel guscio, introducendo enzimi digestivi all'interno del mollusco. Questo processo può indebolire i forti muscoli adduttori (di chiusura) di un bivalve e dare inizio al processo di digestione.
Collegamento all'apprendimento
modificaEsplora da vicino la struttura corporea della stella marina , osservane una muoversi sul fondale marino e osservala mentre divora una cozza.
Le stelle fragili appartengono alla classe Ophiuroidea ("code di serpente"). A differenza delle stelle marine, che hanno braccia paffute, le stelle fragili hanno braccia lunghe, sottili e flessibili che sono nettamente demarcate dal disco centrale. Le stelle fragili si muovono sferzando le braccia o avvolgendole attorno agli oggetti e tirandosi in avanti. Le loro braccia sono anche utilizzate per afferrare le prede. Il sistema vascolare dell'acqua negli ofiuroidi non è utilizzato per la locomozione.
I ricci di mare e i dollari di sabbia sono esempi di Echinoidea ("spinosi"). Questi echinodermi non hanno braccia, ma sono emisferici o appiattiti con cinque file di piedi tubolari che si estendono attraverso cinque file di pori in una conchiglia interna continua chiamata test. I loro piedi tubolari sono usati per mantenere pulita la superficie del corpo. Le placche scheletriche attorno alla bocca sono organizzate in una complessa struttura di alimentazione multiparte chiamata "lanterna di Aristotele ". La maggior parte degli echinoidi bruca le alghe, ma alcuni sono sospensori e altri possono nutrirsi di piccoli animali o detriti organici , i resti frammentari di piante o animali.
I gigli di mare e le stelle piumate sono esempi di Crinoidea. I gigli di mare sono sessili , con il corpo attaccato a uno stelo, ma le stelle piumate possono muoversi attivamente usando cirri simili a zampe che emergono dalla superficie aborale. Entrambi i tipi di crinoide sono sospesi, raccogliendo piccoli organismi alimentari lungo le scanalature ambulacrali dei loro bracci simili a piume. Le "piume" erano costituite da bracci ramificati rivestiti da piedi tubolari. I piedi tubolari vengono utilizzati per spostare il cibo catturato verso la bocca. Esistono solo circa 600 specie di crinoidi esistenti, ma erano molto più numerose e abbondanti negli antichi oceani. Molti crinoidi sono specie di acque profonde, ma le stelle piumate in genere vivono in aree poco profonde, specialmente in acque subtropicali e tropicali.
I cetrioli di mare della classe Holothuroidea presentano un asse orale-aborale esteso. Questi sono gli unici echinodermi che dimostrano una simmetria bilaterale "funzionale" da adulti, perché l'asse orale-aborale esteso costringe l'animale a giacere orizzontalmente piuttosto che stare in verticale. I piedi tubolari sono ridotti o assenti, tranne sul lato su cui giace l'animale. Hanno una singola gonade e il tratto digerente è più tipico di un animale a simmetria bilaterale. Un paio di strutture simili a branchie chiamate alberi respiratori si diramano dall'intestino posteriore; i muscoli attorno alla cloaca pompano acqua dentro e fuori da questi alberi. Ci sono gruppi di tentacoli attorno alla bocca. Alcuni cetrioli di mare si nutrono di detriti, mentre altri sono alimentatori in sospensione, setacciando piccoli organismi con i loro tentacoli orali. Alcune specie di cetrioli di mare sono uniche tra gli echinodermi in quanto le cellule contenenti emoglobina circolano nel fluido celomatico, nel sistema vascolare dell'acqua e/o nel sistema emale.
| a | b | c |
|---|---|---|
 |
 |
 |
| d | e | f |
 |
 |
 |
| Classi di echinodermi. I diversi membri degli Echinodermata includono (a) la stella marina della classe Asteroidea, (b) la stella fragile della classe Ophiuroidea, (c) i ricci di mare della classe Echinoidea, (d) i gigli di mare appartenenti alla classe Crinoidea e (e + f) i cetrioli di mare, che rappresentano la classe Holothuroidea. | ||
Phylum dei Cordati
modificaGli animali del phylum Chordata condividono cinque caratteristiche chiave che compaiono in qualche fase del loro sviluppo: una notocorda, un cordone nervoso cavo dorsale, fessure faringee, una coda post-anale e una ghiandola endostilo/tiroide che secerne ormoni iodati. In alcuni gruppi, alcuni di questi tratti sono presenti solo durante lo sviluppo embrionale. Oltre a contenere classi di vertebrati, il phylum Chordata contiene due cladi di "invertebrati": Urochordata (tunicati, salpe e larve) e Cefalocordati (lancette). La maggior parte dei tunicati vive sul fondale oceanico e si nutre di sospensione. Le lancette sono animali che si nutrono di sospensione e di fitoplancton e altri microrganismi. Gli invertebrati cordati saranno discussi più ampiamente nel capitolo seguente.
Attività
modificaSintesi del capitolo