Biologia per il liceo/I virus
Introduzione
modificaI virus sono entità parassite non cellulari che non possono essere classificate in nessun regno. Possono infettare organismi diversi come batteri, piante e animali. Infatti, i virus esistono in una sorta di mondo sotterraneo tra un organismo vivente e un'entità non vivente. Gli esseri viventi crescono, metabolizzano e si riproducono. Al contrario, i virus non sono cellulari, non hanno un metabolismo o crescono e non possono dividersi per divisione cellulare. I virus possono copiare o replicare se stessi; tuttavia, dipendono interamente dalle risorse derivate dalle loro cellule ospiti per produrre virus progenie, che vengono assemblati nella loro forma matura. Nessuno sa esattamente quando o come i virus si sono evoluti o da quale fonte ancestrale perché i virus non hanno lasciato una documentazione fossile. Alcuni virologi sostengono che i virus moderni sono un mosaico di frammenti di acidi nucleici raccolti da varie fonti lungo i rispettivi percorsi evolutivi. Quasi tutte le forme di vita, dai batteri procarioti e dagli archei, agli eucarioti come piante, animali e funghi, hanno virus che le infettano. Mentre la maggior parte della diversità biologica può essere compresa attraverso la storia evolutiva (ad esempio, come le specie si sono adattate alle mutevoli condizioni ambientali e come le diverse specie sono imparentate tra loro attraverso una discendenza comune), molto sulle origini e l'evoluzione dei virus rimane sconosciuto.
Scoperta e rilevamento
modificaI virus furono scoperti per la prima volta dopo lo sviluppo di un filtro in porcellana, il filtro Chamberland-Pasteur, che poteva rimuovere tutti i batteri visibili al microscopio da qualsiasi campione liquido. Nel 1886, Adolph Meyer dimostrò che una malattia delle piante di tabacco, la malattia del mosaico del tabacco, poteva essere trasferita da una pianta malata a una sana tramite estratti vegetali liquidi. Nel 1892, Dmitri Ivanowski dimostrò che questa malattia poteva essere trasmessa in questo modo anche dopo che il filtro Chamberland-Pasteur aveva rimosso tutti i batteri vitali dall'estratto. Tuttavia, ci vollero molti anni prima che si dimostrasse che questi agenti infettivi "filtrabili" non erano semplicemente batteri molto piccoli, ma un nuovo tipo di particelle molto piccole che causavano malattie.
La maggior parte dei virioni, o singole particelle virali, sono molto piccole, con un diametro di circa 20-250 nanometri. Tuttavia, alcuni virus di amebe scoperti di recente hanno un diametro fino a 1000 nm. Ad eccezione dei grandi virioni, come il poxvirus e altri grandi virus del DNA, i virus non possono essere visti con un microscopio ottico. Solo con lo sviluppo del microscopio elettronico alla fine degli anni '30 gli scienziati hanno ottenuto la prima buona visione della struttura del virus del mosaico del tabacco (TMV) ( vedi galleria), discusso sopra, e di altri virus (vedi galleria ). La struttura superficiale dei virioni può essere osservata sia con la microscopia elettronica a scansione che a trasmissione, mentre le strutture interne del virus possono essere osservate solo in immagini da un microscopio elettronico a trasmissione. L'uso della microscopia elettronica e di altre tecnologie ha permesso la scoperta di molti virus di tutti i tipi di organismi viventi.
-
 Particelle del virus del mosaico del tabacco (TMV) colorate negativamente con metallo pesante per renderle visibili nel TEM. Ingrandimento 160.000X
Particelle del virus del mosaico del tabacco (TMV) colorate negativamente con metallo pesante per renderle visibili nel TEM. Ingrandimento 160.000X -
 Danni del virus del tabacco
Danni del virus del tabacco -
 Foto di foglie di orchidea con sintomi del virus del mosaico del tabacco
Foto di foglie di orchidea con sintomi del virus del mosaico del tabacco -
 Virus batteriofagi che attaccano un batterio
Virus batteriofagi che attaccano un batterio

.jpg)
Evoluzione dei virus
modificaSebbene i biologi abbiano una notevole quantità di conoscenze su come i virus odierni mutano e si adattano, si sa molto meno su come i virus si siano originati in primo luogo. Quando esplorano la storia evolutiva della maggior parte degli organismi, gli scienziati possono guardare ai reperti fossili e a prove storiche simili. Tuttavia, i virus non si fossilizzano, per quanto ne sappiamo, quindi i ricercatori devono estrapolare dalle indagini su come si evolvono i virus odierni e utilizzando informazioni biochimiche e genetiche per creare storie speculative sui virus.
La maggior parte degli studiosi concorda sul fatto che i virus non hanno un singolo antenato comune, né esiste una singola ipotesi ragionevole sulle origini dei virus. Esistono scenari evolutivi attuali che potrebbero spiegare l'origine dei virus. Una di queste ipotesi, la "devolution" o l' ipotesi regressiva , suggerisce che i virus si siano evoluti da cellule viventi libere o da parassiti procariotici intracellulari. Tuttavia, molti componenti di come questo processo potrebbe essere avvenuto rimangono un mistero. Una seconda ipotesi, l' ipotesi escapista o progressiva , suggerisce che i virus abbiano avuto origine da molecole di RNA e DNA, o entità autoreplicanti simili a trasposoni o altri elementi genetici mobili, che sono sfuggite da una cellula ospite con la capacità di entrare in un'altra. Una terza ipotesi, l' ipotesi del primo virus, suggerisce che i virus potrebbero essere state le prime entità autoreplicanti prima delle prime cellule. In tutti i casi, i virus probabilmente continuano a evolversi insieme alle cellule su cui fanno affidamento come ospiti.
Morfologia virale
modificaPer virione si intende la forma completa del virus quando non è dentro una cellula.
I virus sono non cellulari , ovvero entità biologiche prive di struttura cellulare. Pertanto, sono privi della maggior parte dei componenti delle cellule, come organelli, ribosomi e membrana plasmatica. Un virione è costituito da un nucleo di acido nucleico, un rivestimento proteico esterno o capside e talvolta un involucro esterno costituito da membrane proteiche e fosfolipidiche derivate dalla cellula ospite. I virus possono anche contenere proteine aggiuntive, come enzimi, all'interno del capside o attaccate al genoma virale. La differenza più ovvia tra i membri di diverse famiglie virali è la variazione nella loro morfologia, che è piuttosto varia. Una caratteristica interessante della complessità virale è che la complessità dell'ospite non è necessariamente correlata alla complessità del virione. Infatti, alcune delle strutture virioniche più complesse si trovano nei batteriofagi , virus che infettano gli organismi viventi più semplici, i batteri.
Morfologia
modificaI virus hanno molte forme e dimensioni, ma queste caratteristiche sono coerenti per ogni famiglia virale. Come abbiamo visto, tutti i virioni hanno un genoma di acido nucleico coperto da un capside protettivo. Le proteine del capside sono codificate nel genoma virale e sono chiamate capsomeri . Alcuni capsidi virali sono semplici eliche o "sfere" poliedriche, mentre altri hanno una struttura piuttosto complessa (vedi galleria).
-
 Morfologie e dimensioni di diversi virus
Morfologie e dimensioni di diversi virus -
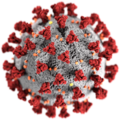 Virus SARS-CoV-2
Virus SARS-CoV-2 -
 Modello schematico del virus batteriofago
Modello schematico del virus batteriofago -
 Modello schematico del virus del mosaico del tabacco
Modello schematico del virus del mosaico del tabacco -
 Esempi di virus a geometria icosaedrica: A. Virus senza membrana; B. Virus con membrana. 1. Capside; 2. Acido nucleico; 3. Capsomero; 4. Nucleocapside; 5. Virione; 6. Rivestimento proteico; 7. Rivestimento glicoproteico.
Esempi di virus a geometria icosaedrica: A. Virus senza membrana; B. Virus con membrana. 1. Capside; 2. Acido nucleico; 3. Capsomero; 4. Nucleocapside; 5. Virione; 6. Rivestimento proteico; 7. Rivestimento glicoproteico. -
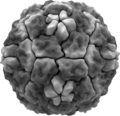 Virus del raffreddore HRV14
Virus del raffreddore HRV14 -
 Struttura e organizzazione genomica del rinovirus umano
Struttura e organizzazione genomica del rinovirus umano -
 Virus del vaiolo
Virus del vaiolo -
 Virione dell'HIV
Virione dell'HIV -
 Virione del Deltalipothrixvirus (infetta particolari batteri archea)
Virione del Deltalipothrixvirus (infetta particolari batteri archea) -
 Virus dell'ebola
Virus dell'ebola -
 Coccolithovirus, infettano dei protisti fotosintetici (alghe)
Coccolithovirus, infettano dei protisti fotosintetici (alghe)











In generale, i capsidi dei virus sono classificati in quattro gruppi: elicoidali, icosaedrici, avvolti e testa e coda. I capsidi elicoidali sono lunghi e cilindrici. Molti virus delle piante sono elicoidali, incluso il TMV. I virus icosaedrici hanno forme approssimativamente sferiche, come quelle del poliovirus o degli herpesvirus. I virus avvolti hanno membrane derivate dalla cellula ospite che circonda i capsidi. I virus animali, come l'HIV, sono spesso avvolti. I virus testa e coda infettano i batteri e hanno una testa simile ai virus icosaedrici e una coda a forma di virus elicoidali.
Molti virus usano una sorta di glicoproteina per attaccarsi alle cellule ospiti tramite molecole sulla cellula chiamate recettori virali . Per questi virus, l'attaccamento è necessario per la successiva penetrazione della membrana cellulare; solo dopo che la penetrazione ha luogo il virus può completare la sua replicazione all'interno della cellula. I recettori che i virus usano sono molecole che si trovano normalmente sulle superfici cellulari e hanno le loro funzioni fisiologiche. Sembra che i virus si siano semplicemente evoluti per utilizzare queste molecole per la loro replicazione. Ad esempio, l'HIV usa la molecola CD4 sui linfociti T come uno dei suoi recettori ( Figura sotto). CD4 è un tipo di molecola chiamata molecola di adesione cellulare , che funziona per mantenere diversi tipi di cellule immunitarie in stretta prossimità l'una all'altra durante la generazione di una risposta immunitaria dei linfociti T.

Uno dei virioni più complessi conosciuti, il batteriofago T4 (che infetta il batterio Escherichia coli ), ha una struttura a coda che il virus usa per attaccarsi alle cellule ospiti e una struttura a testa che ospita il suo DNA.
L'adenovirus, un virus animale non capsulato che causa malattie respiratorie negli esseri umani, utilizza punte di glicoproteina che sporgono dai suoi capsomeri per attaccarsi alle cellule ospiti. I virus non capsulati includono anche quelli che causano la poliomielite (poliovirus), le verruche plantari (papillomavirus) e l'epatite A (virus dell'epatite A).
I virioni con involucro, come il virus dell'influenza, sono costituiti da acido nucleico (RNA nel caso dell'influenza) e proteine del capside circondate da un involucro a doppio strato fosfolipidico che contiene proteine codificate dal virus. Le glicoproteine incorporate nell'involucro virale vengono utilizzate per attaccarsi alle cellule ospiti. Altre proteine dell'involucro sono le proteine della matrice che stabilizzano l'involucro e spesso svolgono un ruolo nell'assemblaggio dei virioni della progenie. Varicella, HIV e parotite sono altri esempi di malattie causate da virus con involucro. A causa della fragilità dell'involucro, i virus non avvolti sono più resistenti ai cambiamenti di temperatura, pH e ad alcuni disinfettanti rispetto ai virus avvolti.
Nel complesso, la forma del virione e la presenza o l'assenza di un involucro ci dicono poco sulla malattia che il virus può causare o sulle specie che potrebbe infettare, ma sono comunque mezzi utili per iniziare la classificazione virale ( Figura 21.5 ).
-
 Batteriofago T4
Batteriofago T4 -
 Adenovirus
Adenovirus -
 Schema del virus dell'influenza
Schema del virus dell'influenza -
 Modalità di infezione del virus dell'influenza
Modalità di infezione del virus dell'influenza




In galleria. I virus possono avere una forma complessa o relativamente semplice. Questa figura mostra tre virioni relativamente complessi: il batteriofago T4, con il suo gruppo di testa contenente DNA e fibre della coda che si attaccano alle cellule ospiti; l'adenovirus, che usa le punte del suo capside per legarsi alle cellule ospiti; e il virus dell'influenza, che usa le glicoproteine incorporate nel suo involucro per legarsi alle cellule ospiti. Il virus dell'influenza ha anche proteine della matrice, interne all'involucro, che aiutano a stabilizzare la forma del virione.
Quale delle seguenti affermazioni sulla struttura del virus è vera?
- Tutti i virus sono racchiusi in una membrana virale.
- Il capsomero è costituito da piccole subunità proteiche chiamate capsidi.
- Il DNA è il materiale genetico presente in tutti i virus.
- Le glicoproteine aiutano il virus ad attaccarsi alla cellula ospite.
Tipi di acido nucleico
modificaA differenza di quasi tutti gli organismi viventi che usano il DNA come materiale genetico, i virus possono usare sia il DNA che l'RNA. Il nucleo del virus contiene il genoma, ovvero il contenuto genetico totale del virus. I genomi virali tendono a essere piccoli, contenenti solo quei geni che codificano proteine che il virus non può ottenere dalla cellula ospite. Questo materiale genetico può essere a singolo o doppio filamento. Può anche essere lineare o circolare. Mentre la maggior parte dei virus contiene un singolo acido nucleico, altri hanno genomi divisi in diversi segmenti. Il genoma dell'RNA del virus dell'influenza è segmentato, il che contribuisce alla sua variabilità e continua evoluzione, e spiega perché è difficile sviluppare un vaccino contro di esso.
Nei virus a DNA, il DNA virale dirige le proteine di replicazione della cellula ospite per sintetizzare nuove copie del genoma virale e per trascrivere e tradurre quel genoma in proteine virali. Le malattie umane causate dai virus a DNA includono la varicella, l'epatite B e gli adenovirus. I virus a DNA trasmessi sessualmente includono l'herpes virus e il papilloma virus umano (HPV), che è stato associato al cancro cervicale e alle verruche genitali.
I virus a RNA contengono solo RNA come materiale genetico. Per replicare i loro genomi nella cellula ospite, i virus a RNA devono codificare i propri enzimi che possono replicare l'RNA in RNA o, nei retrovirus, in DNA. Questi enzimi RNA polimerasi hanno maggiori probabilità di commettere errori di copia rispetto alle DNA polimerasi e quindi spesso commettono errori durante la trascrizione. Per questo motivo, le mutazioni nei virus a RNA si verificano più frequentemente che nei virus a DNA. Ciò fa sì che cambino e si adattino più rapidamente al loro ospite. Le malattie umane causate dai virus a RNA includono l'influenza, l'epatite C, il morbillo e la rabbia. Il virus HIV, che si trasmette sessualmente, è un retrovirus a RNA.
La sfida della classificazione dei virus
modificaPoiché la maggior parte dei virus probabilmente si è evoluta da antenati diversi, i metodi sistematici che gli scienziati hanno utilizzato per classificare le cellule procariotiche ed eucariotiche non sono molto utili. Se i virus rappresentano "resti" di organismi diversi, allora anche l'analisi genomica o proteica non è utile. Perché? Perché i virus non hanno una sequenza genomica comune che condividono tutti . Ad esempio, la sequenza 16S rRNA così utile per costruire filogenesi procariotiche non è di alcuna utilità per una creatura senza ribosomi! I biologi hanno utilizzato diversi sistemi di classificazione in passato. I virus erano inizialmente raggruppati in base alla morfologia condivisa. Successivamente, i gruppi di virus sono stati classificati in base al tipo di acido nucleico che contenevano, DNA o RNA, e se il loro acido nucleico era a singolo o doppio filamento. Tuttavia, questi precedenti metodi di classificazione raggruppavano i virus in modo diverso, perché erano basati su diversi set di caratteri del virus. Il metodo di classificazione più comunemente utilizzato oggi è denominato schema di classificazione di Baltimora e si basa sul modo in cui l'RNA messaggero (mRNA) viene generato in ciascun particolare tipo di virus.
Sistemi di classificazione passati
modificaI virus contengono solo pochi elementi in base ai quali possono essere classificati: il genoma virale, il tipo di capside e la struttura dell'involucro per i virus con involucro. Tutti questi elementi sono stati utilizzati in passato per la classificazione virale (Tabella sotto). I genomi virali possono variare nel tipo di materiale genetico (DNA o RNA) e nella sua organizzazione (a singolo o doppio filamento, lineare o circolare e segmentato o non segmentato). In alcuni virus, proteine aggiuntive necessarie per la replicazione sono associate direttamente al genoma o contenute all'interno del capside virale.
Classificazione dei virus in base alla struttura del genoma
| Struttura del genoma | Esempi | ||
|---|---|---|---|
|
|
 |
 |
|
|
 |
 |
|
|
||
|
|
Tabella sopra. I virus possono essere classificati in base al loro materiale genetico di base e al design del capside. Il virus della rabbia ha un core di RNA a singolo filamento (ssRNA) e un capside elicoidale avvolto, mentre il virus del vaiolo, l'agente causale del vaiolo, ha un core di DNA a doppio filamento (dsDNA) e un capside complesso. La trasmissione della rabbia avviene quando la saliva di un mammifero infetto entra in una ferita. Il virus viaggia attraverso i neuroni nel sistema nervoso periferico fino al sistema nervoso centrale, dove compromette la funzione cerebrale, e poi viaggia verso altri tessuti. Il virus può infettare qualsiasi mammifero e la maggior parte muore entro poche settimane dall'infezione. Il vaiolo è un virus umano trasmesso per inalazione del virus del vaiolo, localizzato nella pelle, nella bocca e nella gola, che causa una caratteristica eruzione cutanea. Prima della sua eradicazione nel 1979, l'infezione determinava un tasso di mortalità del 30-35 percento.
I virus possono anche essere classificati in base al design dei loro capsidi ( Tabella sotto ). I capsidi sono classificati come icosaedrici nudi, icosaedrici avvolti, elicoidali avvolti, elicoidali nudi e complessi. Il tipo di materiale genetico (DNA o RNA) e la sua struttura (a singolo o doppio filamento, lineare o circolare e segmentata o non segmentata) sono utilizzati per classificare le strutture del nucleo del virus.
Classificazione dei virus in base alla struttura del capside
| Classificazione del capside | Esempi | |
|---|---|---|
| Icosaedrico nudo | Virus dell'epatite A, poliovirus |  |
| Icosaedrico avvolto | Virus di Epstein-Barr,
virus herpes simplex, virus della rosolia, virus della febbre gialla, HIV-1 |
 |
| Elicoidale avvolto | Virus influenzali, virus della parotite,
virus del morbillo, virus della rabbia |
 |
| Elicoidale nudo | Virus del mosaico del tabacco |  |
| Complesso con molte proteine; | Herpesvirus, virus del vaiolo,
virus dell'epatite B, batteriofago T4 |
 |
Tabella Le micrografie elettroniche a trasmissione di vari virus mostrano le strutture del loro capside. Il capside del (a) virus della poliomielite è icosaedrico nudo; (b) il capside del virus di Epstein-Barr è icosaedrico avvolto; (c) il capside del virus della parotite è un'elica avvolta; (d) il capside del virus del mosaico del tabacco è elicoidale nudo; e (e) il capside dell'herpesvirus è complesso.
Classificazione di Baltimora
modificaIl sistema di classificazione dei virus più comunemente e attualmente utilizzato è stato sviluppato per la prima volta dal biologo premio Nobel David Baltimore nei primi anni '70. Oltre alle differenze di morfologia e genetica menzionate sopra, lo schema di classificazione di Baltimore raggruppa i virus in base al modo in cui l'mRNA viene prodotto durante il ciclo replicativo del virus.
I virus del gruppo I contengono DNA a doppio filamento (dsDNA) come genoma. Il loro mRNA è prodotto tramite trascrizione in modo molto simile al DNA cellulare, utilizzando gli enzimi della cellula ospite.
I virus del gruppo II hanno DNA a singolo filamento (ssDNA) come genoma. Convertono i loro genomi a singolo filamento in un intermedio dsDNA prima che possa avvenire la trascrizione in mRNA.
I virus del gruppo III usano dsRNA come genoma. I filamenti si separano e uno di essi viene usato come stampo per la generazione di mRNA usando la RNA polimerasi RNA-dipendente codificata dal virus.
I virus del gruppo IV hanno ssRNA come genoma con una polarità positiva , il che significa che l'RNA genomico può fungere direttamente da mRNA. Gli intermedi del dsRNA, chiamati intermedi replicativi , vengono realizzati nel processo di copia dell'RNA genomico. Da questi intermedi si formano molteplici filamenti di RNA a lunghezza intera di polarità negativa (complementari all'RNA genomico a filamento positivo), che possono quindi fungere da modelli per la produzione di RNA con polarità positiva, inclusi sia l'RNA genomico a lunghezza intera che gli mRNA virali più corti.
I virus del gruppo V contengono genomi ssRNA con una polarità negativa , il che significa che la loro sequenza è complementare all'mRNA. Come con i virus del gruppo IV, gli intermedi dsRNA vengono utilizzati per creare copie del genoma e produrre mRNA. In questo caso, il genoma a filamento negativo può essere convertito direttamente in mRNA. Inoltre, i filamenti di RNA positivi a lunghezza intera vengono creati per fungere da stampi per la produzione del genoma a filamento negativo.
I virus del gruppo VI hanno genomi ssRNA diploidi (due copie) che devono essere convertiti, utilizzando l'enzima trascrittasi inversa , in dsDNA; il dsDNA viene quindi trasportato nel nucleo della cellula ospite e inserito nel genoma ospite. Quindi, l'mRNA può essere prodotto tramite la trascrizione del DNA virale che è stato integrato nel genoma ospite.
I virus del gruppo VII hanno genomi dsDNA parziali e producono intermedi ssRNA che agiscono come mRNA, ma vengono anche riconvertiti in genomi dsDNA dalla trascrittasi inversa, necessaria per la replicazione del genoma.
Le caratteristiche di ciascun gruppo nella classificazione di Baltimora sono riassunte nella Tabella sotto con esempi per ciascun gruppo.
Classificazione di Baltimora
| Gruppo | Caratteristiche | Modalità di produzione dell'mRNA | Esempio |
|---|---|---|---|
| IO | DNA a doppio filamento | L'mRNA viene trascritto direttamente dal modello del DNA | Herpes simplex (virus dell'herpes) |
| Io sono | DNA a singolo filamento | Il DNA viene convertito in forma a doppio filamento prima che l'RNA venga trascritto | Parvovirus canino (parvovirus) |
| III | RNA a doppio filamento | L'mRNA viene trascritto dal genoma dell'RNA | Gastroenterite infantile (rotavirus) |
| IV | RNA a singolo filamento (+) | Il genoma funziona come mRNA | Raffreddore comune (picornavirus) |
| V | RNA a singolo filamento (-) | L'mRNA viene trascritto dal genoma dell'RNA | Rabbia (rabdovirus) |
| NOI | Virus a RNA a singolo filamento con trascrittasi inversa | La trascrittasi inversa produce DNA dal genoma dell'RNA; il DNA viene quindi incorporato nel genoma dell'ospite; l'mRNA viene trascritto dal DNA incorporato | Virus dell'immunodeficienza umana (HIV) |
| VII | Virus a DNA a doppio filamento con trascrittasi inversa | Il genoma virale è costituito da DNA a doppio filamento, ma il DNA virale viene replicato attraverso un intermedio RNA; l'RNA può fungere direttamente da mRNA o da stampo per produrre mRNA | Virus dell'epatite B (hepadnavirus) |
Fasi delle infezioni da virus
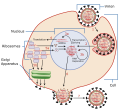
modificaI virus sono parassiti intracellulari obbligati. Un virus deve prima riconoscere e attaccarsi a una specifica cellula vivente prima di entrarvi. Dopo la penetrazione, il virus invasore deve copiare il suo genoma e produrre le proprie proteine. Infine, i virioni della progenie devono sfuggire alla cellula ospite in modo da poter infettare altre cellule. I virus possono infettare solo alcune specie di ospiti e solo alcune cellule all'interno di quell'ospite. Le cellule ospiti specifiche che un virus deve occupare e utilizzare per replicarsi sono chiamate permissive . Nella maggior parte dei casi, la base molecolare di questa specificità è dovuta a una particolare molecola di superficie nota come recettore virale sulla superficie della cellula ospite. È necessario uno specifico recettore virale affinché il virus si attacchi. Inoltre, le differenze nel metabolismo e nelle risposte immunitarie delle cellule ospiti (basate sull'espressione genica differenziale) sono un probabile fattore nel determinare quali cellule un virus può prendere di mira per la replicazione.
Un virus deve usare i suoi processi cellulari ospiti per replicarsi. Il ciclo di replicazione virale può produrre cambiamenti biochimici e strutturali drammatici nella cellula ospite, che possono causare danni cellulari. Questi cambiamenti, chiamati effetti citopatici , possono modificare le funzioni cellulari o persino distruggere la cellula. Alcune cellule infette, come quelle infettate dal comune virus del raffreddore noto come rinovirus, muoiono tramite lisi (scoppio) o apoptosi (morte cellulare programmata o "suicidio cellulare"), rilasciando tutti i virioni della progenie in una volta. I sintomi delle malattie virali derivano sia da tale danno cellulare causato dal virus sia dalla risposta immunitaria al virus, che tenta di controllare ed eliminare il virus dal corpo.
Molti virus animali, come l'HIV (virus dell'immunodeficienza umana) , lasciano le cellule infette del sistema immunitario tramite un processo noto come gemmazione , in cui i virioni lasciano la cellula individualmente. Durante il processo di gemmazione, la cellula non subisce lisi e non viene immediatamente uccisa. Tuttavia, il danno alle cellule che il virus infetta può rendere impossibile il normale funzionamento delle cellule, anche se le cellule rimangono vive per un periodo di tempo. La maggior parte delle infezioni virali produttive segue fasi simili nel ciclo di replicazione del virus: attacco, penetrazione, svestimento, replicazione, assemblaggio e rilascio ( Figura sopra ).
Attacco
modificaUn virus si attacca a uno specifico sito recettore sulla membrana della cellula ospite tramite proteine di attacco nel capside o tramite glicoproteine incorporate nell'involucro virale. La specificità di questa interazione determina l'ospite (e le cellule al suo interno) che possono essere infettate da un virus particolare. Ciò può essere illustrato pensando a diverse chiavi e diverse serrature, dove ogni chiave si adatta solo a una serratura specifica.
Video
modificaQuesto video spiega come l'influenza attacca l'organismo: A Virus Attacks a Cell di Vaccine Makers Project
Ingresso
modificaI virus possono entrare in una cellula ospite con o senza il capside virale. L'acido nucleico dei batteriofagi entra nella cellula ospite "nudo", lasciando il capside all'esterno della cellula. I virus vegetali e animali possono entrare tramite endocitosi (come ricorderai, la membrana cellulare circonda e ingloba l'intero virus). Alcuni virus con involucro entrano nella cellula quando l'involucro virale si fonde direttamente con la membrana cellulare. Una volta all'interno della cellula, il capside virale si degrada e quindi l'acido nucleico virale viene rilasciato e diventa disponibile per la replicazione e la trascrizione.
Replicazione e assemblaggio
modificaIl meccanismo di replicazione dipende dal genoma virale. I virus a DNA solitamente usano proteine ed enzimi della cellula ospite per replicare il DNA virale e trascrivere l'mRNA virale, che viene poi usato per dirigere la sintesi proteica virale. I virus a RNA solitamente usano l'RNA virale come stampo per la sintesi dell'RNA genomico virale e dell'mRNA. L'mRNA virale dirige la cellula ospite a sintetizzare enzimi virali e proteine del capside e ad assemblare nuovi virioni.
Naturalmente, ci sono delle eccezioni a questo schema. Se una cellula ospite non fornisce gli enzimi necessari per la replicazione virale, i geni virali forniscono le informazioni per dirigere la sintesi delle proteine mancanti. I retrovirus, come l'HIV (gruppo VI dello schema di classificazione di Baltimora), hanno un genoma di RNA che deve essere trascritto inversamente in DNA, che viene poi incorporato nel genoma della cellula ospite. Per convertire l'RNA in DNA, i retrovirus devono contenere geni che codificano l'enzima trascrittasi inversa che trascrive un modello di RNA in DNA. La trascrizione inversa non si verifica mai nelle cellule ospiti non infette: l'enzima trascrittasi inversa deriva solo dall'espressione di geni virali all'interno delle cellule ospiti infette. Il fatto che l'HIV produca alcuni dei suoi enzimi non presenti nell'ospite ha permesso ai ricercatori di sviluppare farmaci che inibiscono questi enzimi senza influenzare il metabolismo dell'ospite.
Questo approccio ha portato allo sviluppo di una serie di farmaci utilizzati per curare l'HIV e si è rivelato efficace nel ridurre il numero di virioni infettivi (copie di RNA virale) nel sangue a livelli non rilevabili in molti individui infetti da HIV.
Uscita
modificaL'ultima fase della replicazione virale è il rilascio dei nuovi virioni prodotti nell'organismo ospite, dove sono in grado di infettare le cellule adiacenti e ripetere il ciclo di replicazione. Alcuni virus vengono rilasciati quando la cellula ospite muore, e altri virus possono lasciare le cellule infette gemmando attraverso la membrana senza uccidere direttamente la cellula.

Domanda. Il virus dell'influenza è confezionato in un involucro virale che si fonde con la membrana plasmatica. In questo modo, il virus può uscire dalla cellula ospite senza ucciderla. Quale vantaggio ottiene il virus mantenendo in vita la cellula ospite?
Video
modificaGuarda un video sui virus, sull'identificazione delle strutture, sulle modalità di trasmissione, sulla replicazione e altro ancora.
Batteriofagi
modificaCome hai imparato, i virus spesso infettano ospiti molto specifici, così come cellule specifiche all'interno dell'ospite. Questa caratteristica di un virus lo rende specifico per una o poche specie di vita sulla Terra. D'altra parte, sulla Terra esistono così tanti tipi diversi di virus che quasi ogni organismo vivente ha il suo set di virus che cercano di infettare le sue cellule. Anche i procarioti, le cellule più piccole e semplici, possono essere attaccati da tipi specifici di virus. Nella sezione seguente, esamineremo alcune delle caratteristiche dell'infezione virale delle cellule procariotiche. Come abbiamo imparato, i virus che infettano i batteri sono chiamati batteriofagi ( Figura sotto). Gli archea hanno i loro virus simili.

La maggior parte dei batteriofagi sono virus dsDNA, che utilizzano enzimi dell'ospite per la replicazione del DNA e la trascrizione dell'RNA. Le particelle fagiche devono legarsi a specifici recettori di superficie e inserire attivamente il genoma nella cellula ospite. (Le complesse strutture della coda osservate in molti batteriofagi sono attivamente coinvolte nel far passare il genoma virale attraverso la parete cellulare procariotica.) Quando l'infezione di una cellula da parte di un batteriofago determina la produzione di nuovi virioni, si dice che l'infezione è produttiva . Se i virioni vengono rilasciati facendo scoppiare la cellula, il virus si replica per mezzo di un ciclo litico ( Figura sotto). Un esempio di batteriofago litico è T4, che infetta l'Escherichia coli presente nel tratto intestinale umano. A volte, tuttavia, un virus può rimanere all'interno della cellula senza essere rilasciato. Ad esempio, quando un batteriofago temperato infetta una cellula batterica, si replica per mezzo di un ciclo lisogeno ( Figura sotto) e il genoma virale viene incorporato nel genoma della cellula ospite. Quando il DNA del fago viene incorporato nel genoma della cellula ospite, viene chiamato profago . Un esempio di batteriofago lisogeno è il virus λ (lambda), che infetta anche il batterio E. coli . I virus che infettano le cellule vegetali o animali possono talvolta subire infezioni in cui non producono virioni per lunghi periodi. Un esempio sono gli herpesvirus animali , inclusi i virus herpes simplex, la causa dell'herpes orale e genitale negli esseri umani. In un processo chiamato latenza , questi virus possono esistere nel tessuto nervoso per lunghi periodi di tempo senza produrre nuovi virioni, solo per uscire periodicamente dalla latenza e causare lesioni nella pelle dove il virus si replica. Sebbene vi siano somiglianze tra lisogenia e latenza, il termine ciclo lisogeno è solitamente riservato per descrivere i batteriofagi. La latenza sarà descritta più in dettaglio nella prossima sezione.

Esercizio. Quale delle seguenti affermazioni è falsa?
- Nel ciclo litico vengono prodotti nuovi fagi che vengono rilasciati nell'ambiente.
- Nel ciclo lisogenico, il DNA del fago viene incorporato nel genoma dell'ospite.
- Uno stress ambientale può indurre il fago ad avviare il ciclo lisogeno.
- La lisi cellulare avviene solo nel ciclo litico.
Virus delle piante
modificaLa maggior parte dei virus delle piante, come il virus del mosaico del tabacco, ha genomi di RNA a singolo filamento (+). Tuttavia, ci sono anche virus delle piante nella maggior parte delle altre categorie di virus. A differenza dei batteriofagi, i virus delle piante non hanno meccanismi attivi per trasportare il genoma virale attraverso la parete cellulare protettiva. Affinché un virus delle piante entri in una nuova pianta ospite, deve verificarsi un qualche tipo di danno meccanico. Questo danno è spesso causato da condizioni meteorologiche, insetti, animali, incendi o attività umane come l'agricoltura o la cura del paesaggio. Il movimento da una cellula all'altra all'interno di una pianta può essere facilitato dalla modifica virale dei plasmodesmi (filamenti citoplasmatici che passano da una cellula vegetale all'altra). Inoltre, la prole delle piante può ereditare malattie virali dalle piante madri. I virus delle piante possono essere trasmessi da una varietà di vettori, attraverso il contatto con la linfa di una pianta infetta, da organismi viventi come insetti e nematodi e attraverso il polline. Il trasferimento di un virus da una pianta all'altra è noto come trasmissione orizzontale , mentre l'eredità di un virus da un genitore è chiamata trasmissione verticale .
I sintomi delle malattie virali variano a seconda del virus e del suo ospite ( Tabella 21.4 ). Un sintomo comune è l'iperplasia , la proliferazione anomala di cellule che causa la comparsa di tumori vegetali noti come galle . Altri virus inducono ipoplasia , o diminuzione della crescita cellulare, nelle foglie delle piante, causando la comparsa di aree sottili e gialle. Altri virus ancora colpiscono la pianta uccidendo direttamente le cellule vegetali, un processo noto come necrosi cellulare . Altri sintomi dei virus delle piante includono foglie malformate; striature nere sugli steli delle piante; crescita alterata di steli, foglie o frutti; e macchie ad anello, che sono aree circolari o lineari di scolorimento presenti in una foglia.
Alcuni sintomi comuni delle malattie virali delle piante
| Sintomo | Appare come | |
|---|---|---|
| Iperplasia | Galle (tumori) | _Crown_gall_disease_(49772805266).jpg) |
| Ipoplasia | Macchie gialle e sottili sulle foglie |  |
| Necrosi cellulare | Steli, foglie o frutti morti o anneriti |  |
| Modelli di crescita anormali | Steli, foglie o frutti malformati | _Elizabeth_Bush,_Virginia_Polytechnic_Institute_and_State_University.jpg) |
| Scolorimento | Linee o anelli gialli, rossi
o neri su steli, foglie o frutti |
_-_8529306528.jpg) |
I virus delle piante possono seriamente compromettere la crescita e lo sviluppo delle colture, influenzando significativamente la nostra fornitura alimentare. Sono responsabili della scarsa qualità e quantità delle colture a livello globale e possono causare enormi perdite economiche ogni anno. Altri virus possono danneggiare le piante utilizzate per l'abbellimento del paesaggio. Nelle piante utilizzate per l'abbellimento del paesaggio, due dei virus più comuni sono il virus dell'anulare della peonia e il virus del mosaico della rosa. Ci sono troppi virus delle piante per discuterne in dettaglio, ma i sintomi del virus del mosaico comune del fagiolo comportano una produzione ridotta di fagioli e piante stentate e improduttive. Nella rosa ornamentale, la malattia del mosaico della rosa causa linee gialle ondulate e macchie colorate sulle foglie della pianta.
Virus animali
modificaI virus animali, a differenza dei virus delle piante e dei batteri, non devono penetrare una parete cellulare per accedere alla cellula ospite. Il virus può persino indurre la cellula ospite a collaborare nel processo di infezione. I virus animali non avvolti o "nudi" possono entrare nelle cellule in due modi diversi. Quando una proteina nel capside virale si lega al suo recettore sulla cellula ospite, il virus può essere portato all'interno della cellula tramite una vescicola durante il normale processo cellulare di endocitosi mediata dal recettore . Un metodo alternativo di penetrazione cellulare utilizzato dai virus non avvolti è che le proteine del capside subiscano cambiamenti di forma dopo il legame al recettore, creando canali nella membrana della cellula ospite. Il genoma virale viene quindi "iniettato" nella cellula ospite attraverso questi canali in un modo analogo a quello utilizzato da molti batteriofagi.
I virus con involucro hanno anche due modi per entrare nelle cellule dopo essersi legati ai loro recettori: endocitosi mediata da recettore o fusione . Molti virus con involucro entrano nella cellula tramite endocitosi mediata da recettore in un modo simile a quello visto in alcuni virus senza involucro. D'altro canto, la fusione avviene solo con i virioni con involucro. Questi virus, che includono l'HIV tra gli altri, usano speciali proteine di fusione nei loro involucri per far sì che l'involucro si fonda con la membrana plasmatica della cellula, rilasciando così il genoma e il capside del virus nel citoplasma della cellula.
Dopo aver prodotto le loro proteine e copiato i loro genomi, i virus animali completano l'assemblaggio di nuovi virioni ed escono dalla cellula. Come abbiamo già discusso usando l'esempio del virus dell'influenza, i virus animali con involucro possono gemmare dalla membrana cellulare mentre si assemblano, prendendo un pezzo della membrana plasmatica della cellula nel processo. D'altro canto, la progenie virale non con involucro, come i rinovirus, si accumula nelle cellule infette finché non c'è un segnale per la lisi o l'apoptosi, e tutti i virioni vengono rilasciati insieme.
Come imparerai nel prossimo modulo, i virus animali sono associati a una varietà di malattie umane. Alcune di esse seguono il classico schema della malattia acuta , in cui i sintomi peggiorano progressivamente per un breve periodo, seguito dall'eliminazione del virus dal corpo da parte del sistema immunitario e dall'eventuale guarigione dall'infezione. Esempi di malattie virali acute sono il comune raffreddore e l'influenza. Altri virus causano infezioni croniche a lungo termine , come il virus che causa l'epatite C, mentre altri, come il virus herpes simplex, causano solo sintomi intermittenti . Altri virus ancora, come gli herpesvirus umani 6 e 7, che in alcuni casi possono causare la malattia infantile minore roseola, spesso causano con successo infezioni produttive senza causare alcun sintomo nell'ospite, e quindi diciamo che questi pazienti hanno un'infezione asintomatica .
Nelle infezioni da epatite C, il virus cresce e si riproduce nelle cellule del fegato, causando bassi livelli di danno epatico. Il danno è così basso che gli individui infetti spesso non si rendono conto di esserlo e molte infezioni vengono rilevate solo tramite analisi del sangue di routine su pazienti con fattori di rischio come l'uso di droghe per via endovenosa. D'altro canto, poiché molti dei sintomi delle malattie virali sono causati da risposte immunitarie, la mancanza di sintomi è un'indicazione di una debole risposta immunitaria al virus. Ciò consente al virus di sfuggire all'eliminazione da parte del sistema immunitario e di persistere negli individui per anni, producendo nel contempo bassi livelli di virioni progenitori in quella che è nota come malattia virale cronica. L'infezione cronica del fegato da parte di questo virus porta a una probabilità molto maggiore di sviluppare un cancro al fegato, a volte fino a 30 anni dopo l'infezione iniziale.
Come già discusso, il virus herpes simplex può rimanere in uno stato di latenza nel tessuto nervoso per mesi, persino anni. Poiché il virus si "nasconde" nel tessuto e produce poche proteine virali, se non nessuna, non c'è nulla contro cui la risposta immunitaria possa agire e l'immunità al virus diminuisce lentamente. In determinate condizioni, tra cui vari tipi di stress fisico e psicologico, il virus herpes simplex latente può essere riattivato e subire un ciclo di replicazione litica nella pelle, causando le lesioni associate alla malattia. Una volta che i virioni vengono prodotti nella pelle e le proteine virali vengono sintetizzate, la risposta immunitaria viene nuovamente stimolata e risolve le lesioni cutanee in pochi giorni o settimane distruggendo i virus nella pelle. Come risultato di questo tipo di ciclo replicativo, la comparsa di herpes labiale e focolai di herpes genitale si verificano solo in modo intermittente, anche se i virus rimangono nel tessuto nervoso per tutta la vita. Le infezioni latenti sono comuni anche con altri herpesvirus, tra cui il virus varicella-zoster che causa la varicella. Dopo aver contratto un’infezione da varicella durante l’infanzia, il virus varicella-zoster può rimanere latente per molti anni e riattivarsi negli adulti, causando la dolorosa condizione nota come “fuoco di Sant’Antonio” ( Figura sotto).
_Virus_PHIL_1878_lores.jpg) |
 |
| Un'infezione virale latente. (sinistra) Varicella-zoster, il virus che causa la varicella, ha un capside icosaedrico avvolto visibile in questa micrografia elettronica a trasmissione. Il suo genoma di DNA a doppio filamento viene incorporato nel DNA dell'ospite e può riattivarsi dopo la latenza sotto forma di (destra) herpes zoster, che spesso presenta un'eruzione cutanea | |
Alcuni virus che infettano gli animali, tra cui il virus dell'epatite C discusso sopra, sono noti come virus oncogeni: hanno la capacità di causare il cancro. Questi virus interferiscono con la normale regolazione del ciclo cellulare dell'ospite introducendo geni che stimolano la crescita cellulare non regolata (oncogeni) o interferendo con l'espressione di geni che inibiscono la crescita cellulare. I virus oncogeni possono essere virus a DNA o a RNA. I tumori noti per essere associati a infezioni virali includono il cancro cervicale, causato dal papillomavirus umano (HPV) ( Figura sotto), il cancro al fegato causato dal virus dell'epatite C, la leucemia a cellule T e diversi tipi di linfoma.
_EM.jpg)
Vaccini per la prevenzione
modificaI virus causano una varietà di malattie negli animali, compresi gli esseri umani, che vanno dal comune raffreddore a malattie potenzialmente fatali come la meningite ( Figura sotto). Queste malattie possono essere curate con farmaci antivirali o vaccini; tuttavia, alcuni virus, come l'HIV, sono in grado sia di eludere la risposta immunitaria sia di mutare all'interno dell'organismo ospite per diventare resistenti ai farmaci antivirali.

Il metodo principale per controllare le malattie virali è la vaccinazione , che ha lo scopo di prevenire le epidemie creando immunità a un virus o a una famiglia di virus ( Figura sotto). I vaccini possono essere preparati utilizzando virus vivi, virus uccisi o subunità molecolari del virus. Si noti che i vaccini virali uccisi e i virus subunitari sono entrambi incapaci di causare malattie, né vi è alcuna prova valida che le vaccinazioni contribuiscano all'autismo.
I vaccini virali vivi sono progettati in laboratorio per causare pochi sintomi nei riceventi, conferendo loro al contempo un'immunità protettiva contro future infezioni. La poliomielite è stata una malattia che ha rappresentato una pietra miliare nell'uso dei vaccini. Le epidemie di poliomielite si sono verificate con frequenza e impatto crescenti con l'avanzare del ventesimo secolo, diventando un evento terrificante e tragico ogni estate. Decine di migliaia di persone sono morte e molte altre sono rimaste paralizzate; i bambini costituivano una grande parte delle vittime. Utilizzando virus uccisi testati sulla linea cellulare HeLa (originariamente ottenuta da Henrietta Lacks e poi prodotta in serie per soddisfare la necessità), Jonas Salk ha sviluppato un vaccino di successo. Le campagne di immunizzazione di massa negli anni '50 (vaccino ucciso) e '60 (vaccino vivo) hanno ridotto significativamente l'incidenza della malattia. Il successo del vaccino contro la poliomielite ha aperto la strada alla distribuzione di routine di vaccini infantili contro morbillo, parotite, rosolia, varicella e altre malattie.

Il problema con l'uso di vaccini vivi (che sono solitamente più efficaci dei vaccini uccisi), è il basso ma significativo pericolo che questi virus tornino alla loro forma patogena tramite retromutazioni . I vaccini vivi sono solitamente realizzati attenuando (indebolendo) il virus "selvatico" (patogeno) facendolo crescere in laboratorio in tessuti o a temperature diverse da quelle a cui il virus è abituato nell'ospite. Gli adattamenti a queste nuove cellule o temperature inducono mutazioni nei genomi del virus, consentendogli di crescere meglio in laboratorio e inibendo la sua capacità di causare malattie quando reintrodotto nelle condizioni riscontrate nell'ospite. Questi virus attenuati causano quindi ancora infezioni, ma non crescono molto bene, consentendo alla risposta immunitaria di svilupparsi in tempo per prevenire malattie gravi. Le retromutazioni si verificano quando il vaccino subisce mutazioni nell'ospite tali da riadattarsi all'ospite e può nuovamente causare malattie, che possono quindi essere trasmesse ad altri esseri umani in un'epidemia. Uno scenario simile si è verificato di recente in Nigeria nel 2007, quando alcune mutazioni in un vaccino contro la poliomielite hanno causato un'epidemia di poliomielite nel Paese.
Alcuni vaccini sono in continuo sviluppo perché certi virus, come l'influenza e l'HIV, hanno un alto tasso di mutazione rispetto a quello di altri virus e normali cellule ospiti. Con l'influenza, le mutazioni nelle molecole di superficie del virus aiutano l'organismo a eludere l'immunità protettiva che potrebbe essere stata ottenuta in una precedente stagione influenzale, rendendo necessario che gli individui si vaccinino ogni anno. Altri virus, come quelli che causano le malattie infantili morbillo, parotite e rosolia, mutano così raramente che lo stesso vaccino viene utilizzato anno dopo anno.
Vaccini e farmaci antivirali per il trattamento
modificaVaccini come terapia
modifica. In alcuni casi, i vaccini possono essere utilizzati per trattare un'infezione virale attiva. Il concetto alla base di ciò è che somministrando il vaccino, l'immunità viene rafforzata senza aggiungere altro virus patogeno. Nel caso della rabbia , una malattia neurologica mortale trasmessa tramite la saliva di animali infetti dal virus della rabbia, la progressione della malattia dal momento del morso dell'animale al momento in cui entra nel sistema nervoso centrale può essere di due settimane o più. Questo è un lasso di tempo sufficiente per vaccinare gli individui che sospettano di essere stati morsi da un animale rabbioso e la loro risposta immunitaria rafforzata è sufficiente a impedire al virus di entrare nel tessuto nervoso. Pertanto, le conseguenze neurologiche potenzialmente fatali della malattia vengono scongiurate e l'individuo deve solo riprendersi dal morso infetto. Questo approccio viene utilizzato anche per il trattamento dell'Ebola, uno dei virus più rapidi e mortali sulla Terra. Trasmessa da pipistrelli e grandi scimmie, questa malattia può causare la morte nel 70-90 percento degli esseri umani infetti entro due settimane. Utilizzando vaccini di nuova concezione che potenziano la risposta immunitaria in questo modo, c'è la speranza che gli individui colpiti saranno in grado di controllare meglio il virus, salvando potenzialmente una percentuale maggiore di persone infette da una morte rapida e molto dolorosa.
Farmaci antivirali
modificaUn altro modo per trattare le infezioni virali è l'uso di farmaci antivirali. Poiché i virus utilizzano le risorse della cellula ospite per la replicazione e la produzione di nuove proteine virali, è difficile bloccare le loro attività senza danneggiare l'ospite. Per molti anni, gli scienziati hanno pensato che i farmaci in grado di avere un impatto sui virus sarebbero stati troppo tossici per l'organismo. Per affrontare questa sfida, la ricercatrice Gertrude Elion ha cercato di sviluppare farmaci che avrebbero preso di mira solo il virus attraverso processi come l'inibizione della sola replicazione del DNA virale. Ad esempio, alcuni dei suoi farmaci si concentravano sulle purine, mentre altri influenzavano la DNA polimerasi. (Elion ha 45 brevetti che spaziano dagli antivirali agli immunosoppressori ai farmaci contro il cancro.) Tuttavia, abbiamo alcuni farmaci antivirali efficaci, come quelli usati per curare l'HIV e l'influenza. Alcuni farmaci antivirali sono specifici per un virus particolare e altri sono stati usati per controllare e ridurre i sintomi di un'ampia varietà di malattie virali. Per la maggior parte dei virus, i farmaci antivirali possono inibire il virus bloccando le azioni di una o più delle sue proteine. È importante notare che le proteine mirate sono codificate da geni virali e che queste molecole non sono presenti in una cellula ospite sana. In questo modo, la crescita virale viene inibita senza danneggiare l'ospite. Il lavoro di Elion con George Hitchens non solo ha portato a trattamenti diretti, ma, cosa più importante, ha cambiato l'intera metodologia di sviluppo dei farmaci. Prendendo di mira aspetti specifici delle cellule tumorali, dei virus e dei batteri, hanno gettato le basi per molti dei farmaci più comuni e importanti di oggi, utilizzati per aiutare milioni di persone ogni anno. Hanno ricevuto il premio Nobel nel 1988.
Sono stati sviluppati antivirali per trattare l'herpes genitale (herpes simplex II) e l'influenza. Per l'herpes genitale, farmaci come l'aciclovir, sviluppato da Elion, possono ridurre il numero e la durata degli episodi di malattia virale attiva, durante i quali i pazienti sviluppano lesioni virali nelle cellule della pelle. Poiché il virus rimane latente nel tessuto nervoso del corpo per tutta la vita, questo farmaco non è curativo, ma può rendere i sintomi della malattia più gestibili. Per l'influenza, farmaci come il Tamiflu (oseltamivir) ( Figura sotto) possono ridurre la durata dei sintomi "influenzali" di uno o due giorni, ma il farmaco non previene completamente i sintomi. Il Tamiflu agisce inibendo un enzima (neuraminidasi virale) che consente ai nuovi virioni di lasciare le cellule infette. Pertanto, il Tamiflu inibisce la diffusione del virus dalle cellule infette a quelle non infette. Altri farmaci antivirali, come la ribavirina, sono stati utilizzati per trattare una varietà di infezioni virali, sebbene il suo meccanismo d'azione contro alcuni virus rimanga poco chiaro.

Di gran lunga, l'uso di antivirali di maggior successo è stato nel trattamento del retrovirus HIV, che causa una malattia che, se non curata, è solitamente fatale entro 10-12 anni dall'infezione. I farmaci anti-HIV sono stati in grado di controllare la replicazione virale al punto che gli individui che hanno ricevuto questi farmaci sopravvivono per un tempo significativamente più lungo rispetto a quelli non curati.
Una sfida particolare con l'HIV è la sua tendenza a mutare rapidamente all'interno del corpo di un singolo paziente. Ciò porta a una resistenza individuale ai farmaci e richiede una strategia di trattamento diversa rispetto a molte altre malattie. David Ho è stato tra i primi a proporre e sviluppare un metodo per trattare più mutazioni dell'HIV contemporaneamente. Gli sforzi di Ho hanno rappresentato una svolta nella lotta all'AIDS. I farmaci anti-HIV inibiscono la replicazione virale in molte fasi diverse del ciclo replicativo dell'HIV ( Figura sotto). Sono stati sviluppati farmaci che inibiscono la fusione dell'involucro virale dell'HIV con la membrana plasmatica della cellula ospite (inibitori della fusione), la conversione del suo genoma RNA in DNA a doppio filamento (inibitori della trascrittasi inversa, come l'AZT), l'integrazione del DNA virale nel genoma ospite (inibitori dell'integrasi) e l'elaborazione delle proteine virali (inibitori della proteasi).

Sfortunatamente, quando uno qualsiasi di questi farmaci viene utilizzato singolarmente, l'elevato tasso di mutazione del virus gli consente di sviluppare facilmente e rapidamente una resistenza al farmaco, limitandone l'efficacia. La svolta nel trattamento dell'HIV è stata lo sviluppo dell'HAART, terapia antiretrovirale altamente attiva , che prevede una miscela di farmaci diversi, a volte chiamata "cocktail" di farmaci. Attaccando il virus in diverse fasi del suo ciclo replicativo, è molto più difficile per il virus sviluppare resistenza a più farmaci contemporaneamente. Tuttavia, anche con l'uso della terapia HAART combinata, c'è il timore che, nel tempo, il virus sviluppi resistenza a questa terapia. Pertanto, vengono costantemente sviluppati nuovi farmaci anti-HIV con la speranza di continuare la battaglia contro questo virus altamente letale.
-
 Ciclo riproduttivo dell'HIV
Ciclo riproduttivo dell'HIV
Altre entità acellulari: prioni e viroidi
modificaI prioni e i viroidi sono agenti patogeni (agenti in grado di causare malattie) dotati di strutture più semplici dei virus ma, nel caso dei prioni, possono comunque provocare malattie mortali.
Prioni
modificaI prioni , così chiamati perché sono proteici, sono particelle infettive, più piccole dei virus, che non contengono acidi nucleici (né DNA né RNA). Storicamente, l'idea di un agente infettivo che non utilizzasse acidi nucleici era considerata impossibile, ma il lavoro pionieristico del biologo premio Nobel Stanley Prusiner ha convinto la maggior parte dei biologi che tali agenti esistono davvero.
Malattie neurodegenerative fatali, come il kuru negli esseri umani e l'encefalopatia spongiforme bovina (BSE) nei bovini (comunemente nota come "mucca pazza"), sono state trasmesse dai prioni. La malattia si diffondeva tramite il consumo di carne, tessuto nervoso o organi interni tra membri della stessa specie. Il kuru, originario degli esseri umani in Papua Nuova Guinea, si diffondeva da uomo a uomo tramite cannibalismo rituale. La BSE, originariamente rilevata nel Regno Unito, si diffondeva tra i bovini tramite la pratica di includere tessuto nervoso bovino nel mangime per altri bovini. Gli individui con kuru e BSE mostrano sintomi di perdita del controllo motorio e comportamenti insoliti, come scoppi di risate incontrollati con il kuru, seguiti dalla morte. Il kuru veniva controllato inducendo la popolazione ad abbandonare il suo cannibalismo rituale.
D'altro canto, inizialmente si pensava che la BSE colpisse solo i bovini. È stato dimostrato che i bovini che morivano di questa malattia avevano sviluppato lesioni o "buchi" nel cervello, che facevano sì che il tessuto cerebrale assomigliasse a una spugna. Più avanti durante l'epidemia, tuttavia, è stato dimostrato che un'encefalopatia simile negli esseri umani, nota come variante della malattia di Creutzfeldt-Jakob (CJD), poteva essere contratta mangiando carne di animali infetti da BSE, innescando divieti da parte di vari paesi sull'importazione di carne bovina britannica e causando notevoli danni economici all'industria della carne bovina britannica ( Figura sotto). La BSE esiste ancora in varie aree e, sebbene sia una malattia rara, gli individui che contraggono la CJD sono difficili da curare. La malattia può essere trasmessa da uomo a uomo tramite il sangue, quindi molti paesi hanno vietato la donazione di sangue dalle regioni associate alla BSE.
La causa delle encefalopatie spongiformi, come il kuru e la BSE, è una variante strutturale infettiva di una normale proteina cellulare chiamata PrP (proteina prionica). È questa variante che costituisce la particella prionica. La PrP esiste in due forme, PrP c , la forma normale della proteina, e PrP sc , la forma infettiva. Una volta introdotta nel corpo, la PrP sc contenuta nel prione si lega alla PrP c e la converte in PrP sc . Ciò porta a un aumento esponenziale della proteina PrP sc , che si aggrega. La PrP sc è ripiegata in modo anomalo e la conformazione (forma) risultante è direttamente responsabile delle lesioni osservate nei cervelli dei bovini infetti. Pertanto, sebbene non senza alcuni detrattori tra gli scienziati, il prione sembra essere probabilmente una forma completamente nuova di agente infettivo, la prima trovata la cui trasmissione non dipende da geni costituiti da DNA o RNA.

Viroidi
modificaI viroidi sono patogeni delle piante: piccole particelle di RNA circolari a singolo filamento, molto più semplici di un virus. Non hanno un capside o un involucro esterno, ma come i virus possono riprodursi solo all'interno di una cellula ospite. I viroidi, tuttavia, non producono proteine e producono solo una singola molecola di RNA specifica. Le malattie umane causate dai viroidi devono ancora essere identificate.
I viroidi sono noti per infettare le piante ( Figura sotto) e sono responsabili di fallimenti nei raccolti e della perdita di milioni di dollari di entrate agricole ogni anno. Alcune delle piante che infettano includono patate, cetrioli, pomodori, crisantemi, avocado e palme da cocco.

-
 Viroidi della familia Avsunviroidae.
Viroidi della familia Avsunviroidae. -
 Viroidi della familia Pospiviroidae.
Viroidi della familia Pospiviroidae. -
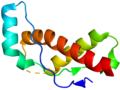 Esempio di prione
Esempio di prione -
 propagazione di un prione
propagazione di un prione -
 Modelli di forme normali (PrPC ) e infettive (PrPSc ) della proteina prionica su una membrana
Modelli di forme normali (PrPC ) e infettive (PrPSc ) della proteina prionica su una membrana -
 Questa mucca affetta da BSE presenta una postura anomala e perdita di peso.
Questa mucca affetta da BSE presenta una postura anomala e perdita di peso.




