Biologia per il liceo/Il ciclo cellulare
Un essere umano, come ogni organismo che si riproduce sessualmente, inizia la vita come un ovulo fecondato (embrione) o zigote . Nella nostra specie, miliardi di divisioni cellulari devono successivamente verificarsi in modo controllato per produrre un essere umano complesso e multicellulare composto da trilioni di cellule. Quindi, lo zigote unicellulare originale è letteralmente l'antenato di tutte le cellule del corpo. Tuttavia, una volta che un essere umano è completamente cresciuto, la riproduzione cellulare è ancora necessaria per riparare e rigenerare i tessuti e talvolta per aumentare le nostre dimensioni! Infatti, tutti gli organismi multicellulari utilizzano la divisione cellulare per la crescita e il mantenimento e la riparazione di cellule e tessuti. La divisione cellulare è strettamente regolata e l'occasionale fallimento di questa regolazione può avere conseguenze pericolose per la vita. Gli organismi unicellulari possono anche utilizzare la divisione cellulare come metodo di riproduzione.
DNA, geni e cromosomi
modificaLa continuità della vita da una cellula all'altra ha il suo fondamento nella riproduzione delle cellule tramite il ciclo cellulare. Il ciclo cellulare è una sequenza ordinata di eventi che descrive le fasi della vita di una cellula dalla divisione di una singola cellula madre alla produzione di due nuove cellule figlie geneticamente identiche.
DNA genomico
modificaPrima di discutere i passaggi che una cellula deve intraprendere per replicare e dividere il suo DNA, è necessaria una comprensione più approfondita della struttura e della funzione delle informazioni genetiche di una cellula. Il DNA di una cellula, confezionato come una molecola di DNA a doppio filamento, è chiamato genoma . Nei procarioti, il genoma è composto da una singola molecola di DNA a doppio filamento a forma di anello o cerchio ( Figura sotto, in rosso). La regione nella cellula che contiene questo materiale genetico è chiamata nucleoide. Alcuni procarioti hanno anche anelli di DNA più piccoli chiamati plasmidi che non sono essenziali per la normale crescita. I batteri possono scambiare questi plasmidi con altri batteri, a volte ricevendo nuovi geni benefici che il ricevente può aggiungere al proprio DNA cromosomico. La resistenza agli antibiotici è una caratteristica che spesso si diffonde attraverso una colonia batterica tramite lo scambio di plasmidi da donatori resistenti a cellule riceventi.
.svg)
Negli eucarioti, il genoma è costituito da diverse molecole di DNA lineare a doppio filamento ( Figura 10.3 ). Ogni specie di eucarioti ha un numero caratteristico di cromosomi nei nuclei delle sue cellule. Le cellule del corpo umano (somatiche) hanno 46 cromosomi, mentre i gameti umani (spermatozoi o ovuli) hanno 23 cromosomi ciascuno. Una tipica cellula del corpo contiene due set di cromosomi corrispondenti o omologhi (un set da ciascun genitore biologico), una configurazione nota come diploide . (Nota: la lettera n è usata per rappresentare un singolo set di cromosomi; pertanto, un organismo diploide è designato 2 n .) Le cellule umane che contengono un set di cromosomi sono chiamate gameti, o cellule sessuali; si tratta di ovuli e spermatozoi, e sono designate 1n , o aploide .
-
 Cromosoma (duplicato)
Cromosoma (duplicato) -
 Struttura del cromosoma
Struttura del cromosoma -
 Cromosoma 17 umano (non duplicato)
Cromosoma 17 umano (non duplicato) -
 Tipica cellula procariote (batteri) con DNA circolare e plasmidi
Tipica cellula procariote (batteri) con DNA circolare e plasmidi -

-
 DNA circolare e plasmidi nella cellula procariote (batteri)
DNA circolare e plasmidi nella cellula procariote (batteri) -
 Struttura della cromatina
Struttura della cromatina -
 Cromosomi omologhi (non duplicati e duplicati)
Cromosomi omologhi (non duplicati e duplicati)






Dopo la fecondazione, ogni gamete contribuisce con un set di cromosomi, creando una cellula diploide contenente coppie di cromosomi corrispondenti chiamati cromosomi omologhi ( "stessa conoscenza") . I cromosomi omologhi hanno la stessa lunghezza e hanno specifici segmenti di nucleotidi chiamati geni esattamente nella stessa posizione, o locus . I geni, le unità funzionali dei cromosomi, determinano caratteristiche specifiche codificando proteine specifiche. I tratti sono le variazioni di tali caratteristiche. Ad esempio, il colore dei capelli è una caratteristica con tratti che sono biondo, castano o nero e molti colori intermedi.

Ogni copia di una coppia omologa di cromosomi ha origine da un genitore diverso; pertanto, i diversi geni (alleli) non sono identici, sebbene codifichino per gli stessi tratti come il "colore dei capelli". La variazione degli individui all'interno di una specie è dovuta alla combinazione specifica dei geni ereditati da entrambi i genitori. Anche una sequenza leggermente alterata di nucleotidi all'interno di un gene può dare origine a un tratto alternativo. Ad esempio, ci sono tre possibili sequenze geniche sul cromosoma umano che codificano per il gruppo sanguigno: sequenza A, sequenza B e sequenza O. Poiché tutte le cellule umane diploidi hanno due copie del cromosoma che determina il gruppo sanguigno, il gruppo sanguigno (il tratto) è determinato dai due alleli del gene marcatore che vengono ereditati. È possibile avere due copie della stessa sequenza genica su entrambi i cromosomi omologhi, con una su ciascuno (ad esempio, AA, BB o OO), o due sequenze diverse, come AB, AO o BO.
Variazioni apparentemente minori di tratti, come il gruppo sanguigno, il colore degli occhi e la manualità, contribuiscono alla variazione naturale riscontrata all'interno di una specie, ma anche se sembrano minori, questi tratti possono essere collegati all'espressione di altri tratti ancora sconosciuti. Tuttavia, se si confronta l'intera sequenza di DNA di una qualsiasi coppia di cromosomi omologhi umani, la differenza è molto inferiore all'uno percento. I cromosomi sessuali, X e Y, sono l'unica eccezione alla regola dell'uniformità dei cromosomi omologhi: a parte una piccola quantità di omologia necessaria per produrre accuratamente i gameti, i geni trovati sui cromosomi X e Y sono diversi.
Struttura e compattazione cromosomica eucariotica
modificaSe il DNA di tutti i 46 cromosomi nel nucleo di una cellula umana fosse disposto uno dietro l'altro, misurerebbe circa due metri; tuttavia, il suo diametro sarebbe di soli 2 nm! Considerando che la dimensione di una tipica cellula umana è di circa 10 µm (100.000 cellule allineate equivalgono a un metro), il DNA deve essere strettamente impacchettato per adattarsi al nucleo della cellula. Allo stesso tempo, deve anche essere facilmente accessibile affinché i geni vengano espressi. Per questo motivo, i lunghi filamenti di DNA vengono condensati in cromosomi compatti durante alcune fasi del ciclo cellulare. Esistono diversi modi in cui i cromosomi vengono compattati.
Nel primo livello di compattazione, brevi tratti della doppia elica del DNA si avvolgono attorno a un nucleo di otto proteine istoniche a intervalli regolari lungo l'intera lunghezza del cromosoma (Figura 10.4). Il complesso DNA-istone è chiamato cromatina. Il complesso istonico del DNA a forma di perla è chiamato nucleosoma e il DNA che collega i nucleosomi è chiamato DNA linker. Una molecola di DNA in questa forma è circa sette volte più corta della doppia elica senza gli istoni e le perle hanno un diametro di circa 10 nm, in contrasto con il diametro di 2 nm di una doppia elica del DNA.
Il secondo livello di compattazione avviene quando i nucleosomi e il DNA di collegamento tra di loro si avvolgono in una fibra di cromatina da 30 nm. Questo avvolgimento condensa ulteriormente il cromosoma, che ora è circa 50 volte più corto della forma estesa.
Nel terzo livello di compattazione, una varietà di proteine fibrose viene utilizzata per "impacchettare la cromatina". Queste proteine fibrose assicurano anche che ogni cromosoma in una cellula non in divisione occupi un'area particolare del nucleo che non si sovrapponga a quella di nessun altro cromosoma (vedere l'immagine in alto nella Figura 10.3 ).

Il DNA si replica nella fase S dell'interfase, che tecnicamente non fa parte della mitosi, ma deve sempre precederla . Dopo la replicazione, i cromosomi sono composti da due cromatidi fratelli collegati . Quando sono completamente compatti, le coppie di cromosomi impacchettati in modo identico sono legati tra loro da proteine coesine. La connessione tra i cromatidi fratelli è più stretta in una regione chiamata centromero . I cromatidi fratelli congiunti, con un diametro di circa 1 µm, sono visibili al microscopio ottico. La regione centromerica è altamente condensata e quindi apparirà come un'area ristretta.
Il ciclo cellulare
modificaIl ciclo cellulare è una serie ordinata di eventi che coinvolgono la crescita cellulare e la divisione cellulare che produce due nuove cellule figlie. Le cellule sul percorso verso la divisione cellulare procedono attraverso una serie di fasi di crescita, replicazione del DNA e divisione nucleare e citoplasmatica, programmate con precisione e attentamente regolate, che alla fine producono due cellule identiche (cloni). Il ciclo cellulare ha due fasi principali: interfase e fase mitotica ( Figura sotto). Durante l' interfase , la cellula cresce e il DNA viene replicato. Durante la fase mitotica , il DNA replicato e il contenuto citoplasmatico vengono separati e il citoplasma cellulare viene tipicamente suddiviso da un terzo processo del ciclo cellulare chiamato citochinesi . Tuttavia, dovremmo notare che l'interfase e la mitosi (cariochinesi) possono aver luogo senza citochinesi, nel qual caso vengono prodotte cellule con più nuclei (cellule multinucleate).

Interfase
modificaDurante l'interfase, la cellula subisce normali processi di crescita, preparandosi anche alla divisione cellulare. Affinché una cellula passi dall'interfase alla fase mitotica, devono essere soddisfatte molte condizioni interne ed esterne. Le tre fasi dell'interfase sono chiamate G 1 , S e G 2 .
Fase G 1 (Primo Gap)
modificaLa prima fase dell'interfase è chiamata fase G 1 (primo gap) perché, da un punto di vista microscopico, sono visibili pochi cambiamenti. Tuttavia, durante la fase G 1 , la cellula è piuttosto attiva a livello biochimico. La cellula accumula i mattoni del DNA cromosomico e le proteine associate, oltre ad accumulare sufficienti riserve di energia per completare il compito di replicare ogni cromosoma nel nucleo.
Fase S (Sintesi del DNA)
modificaDurante l'interfase, il DNA nucleare rimane in una configurazione di cromatina semi-condensata. Nella fase S , la replicazione del DNA può procedere attraverso i meccanismi che determinano la formazione di coppie identiche di molecole di DNA, cromatidi fratelli, che sono saldamente attaccate alla regione centromerica. Il centrosoma viene anche duplicato durante la fase S. I due centrosomi daranno origine al fuso mitotico , l'apparato che orchestra il movimento dei cromosomi durante la mitosi. Ad esempio, più o meno al centro di ogni cellula animale, i centrosomi sono associati a una coppia di oggetti simili a bastoncelli, i centrioli , che sono posizionati ad angolo retto l'uno rispetto all'altro. I centrioli aiutano a organizzare la divisione cellulare. Dovremmo notare, tuttavia, che i centrioli non sono presenti nei centrosomi di altri organismi eucariotici, come le piante e la maggior parte dei funghi.
Fase G 2 (Secondo Gap)
modificaNella fase G 2 , la cellula ripristina le sue riserve di energia e sintetizza le proteine necessarie per la manipolazione e il movimento dei cromosomi. Alcuni organelli cellulari vengono duplicati e il citoscheletro viene smantellato per fornire risorse per la fase mitotica. Potrebbe esserci un'ulteriore crescita cellulare durante G 2 . I preparativi finali per la fase mitotica devono essere completati prima che la cellula sia in grado di entrare nella prima fase della mitosi.
La fase mitotica
modificaLa fase mitotica è un processo multifase durante il quale i cromosomi duplicati vengono allineati, separati e spostati in due nuove cellule figlie identiche. La prima parte della fase mitotica è chiamata mitosi, o cariocinesi. Come abbiamo appena visto, la seconda parte della fase mitotica (e spesso vista come un processo separato e successivo alla mitosi) è chiamata citochinesi, ovvero la separazione fisica dei componenti citoplasmatici nelle due cellule figlie.
Mitosi (cariocinesi)
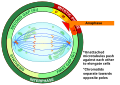
modificaLa cariocinesi, nota anche come mitosi , è divisa in una serie di fasi (profase, prometafase, metafase, anafase e telofase) che portano alla divisione del nucleo cellulare ( Figura sotto).

| Esercizio |
|---|
Quale dei seguenti è l'ordine corretto degli eventi nella mitosi?
|
Profase (la "prima fase"): l'involucro nucleare inizia a dissociarsi in piccole vescicole e gli organelli membranosi (come il complesso del Golgi [apparato del Golgi] e il reticolo endoplasmatico) si frammentano e si disperdono verso la periferia della cellula. Anche il nucleolo scompare (si disperde) e i centrosomi iniziano a spostarsi verso i poli opposti della cellula. I microtubuli che formeranno il fuso mitotico si estendono tra i centrosomi, allontanandoli ulteriormente man mano che le fibre dei microtubuli si allungano. I cromatidi fratelli iniziano ad avvolgersi più strettamente con l'aiuto delle proteine condensina e ora diventano visibili al microscopio ottico.
Prometafase (la "prima fase di cambiamento"): molti processi iniziati nella profase continuano ad avanzare. I resti dell'involucro nucleare si frammentano ulteriormente e il fuso mitotico continua a svilupparsi man mano che più microtubuli si assemblano e si allungano lungo la lunghezza dell'ex area nucleare. I cromosomi diventano ancora più condensati e discreti. Ogni cromatide fratello sviluppa una struttura proteica chiamata cinetocore nella sua regione centromerica ( Figura sotto). Le proteine del cinetocore attraggono e si legano ai microtubuli del fuso mitotico. Quando i microtubuli del fuso si estendono dai centrosomi, alcuni di questi microtubuli entrano in contatto e si legano saldamente ai cinetocori. Una volta che una fibra mitotica si attacca a un cromosoma, il cromosoma sarà orientato finché i cinetocori dei cromatidi fratelli non saranno rivolti verso i poli opposti . Alla fine, tutti i cromatidi fratelli saranno attaccati tramite i loro cinetocori ai microtubuli dai poli opposti. I microtubuli del fuso che non si agganciano ai cromosomi sono chiamati microtubuli polari . Questi microtubuli si sovrappongono a metà strada tra i due poli e contribuiscono all'allungamento cellulare . I microtubuli astrali sono situati vicino ai poli, aiutano nell'orientamento del fuso e sono necessari per la regolazione della mitosi.

Metafase (la "fase di cambiamento"): tutti i cromosomi sono allineati in un piano chiamato piastra metafasica , o piano equatoriale, più o meno a metà strada tra i due poli della cellula. I cromatidi fratelli sono ancora strettamente attaccati l'uno all'altro dalle proteine coesine. In questo momento, i cromosomi sono massimamente condensati.

Anafase ("fase ascendente"): le proteine della coesina si degradano e i cromatidi fratelli si separano al centromero. Ogni cromatide, ora chiamato singolo cromosoma, viene tirato rapidamente verso il centrosoma a cui è attaccato il suo microtubulo. La cellula diventa visibilmente allungata (di forma ovale) mentre i microtubuli polari scivolano l'uno contro l'altro alla piastra metafasica dove si sovrappongono.

Telofase (la "fase di distanza"): i cromosomi raggiungono i poli opposti e iniziano a decondendersi (srotolarsi), rilassandosi ancora una volta in una configurazione di cromatina allungata. I fusi mitotici vengono depolimerizzati in monomeri di tubulina che saranno utilizzati per assemblare i componenti del citoscheletro per ogni cellula figlia. Gli involucri nucleari si formano attorno ai cromosomi e i nucleosomi compaiono all'interno dell'area nucleare.

Citochinesi (citodieresi)
modificaLa citochinesi , o "movimento cellulare", è talvolta considerata la seconda fase principale della fase mitotica, durante la quale la divisione cellulare viene completata tramite la separazione fisica dei componenti citoplasmatici in due cellule figlie. Tuttavia, come abbiamo visto in precedenza, la citochinesi può anche essere considerata una fase separata, che può o meno aver luogo dopo la mitosi. Se la citochinesi ha luogo, la divisione cellulare non è completa finché i componenti cellulari non sono stati suddivisi e completamente separati nelle due cellule figlie. Sebbene le fasi della mitosi siano simili per la maggior parte degli eucarioti, il processo di citochinesi è piuttosto diverso per gli eucarioti che hanno pareti cellulari, come le cellule vegetali.
Nelle cellule animali, la citochinesi inizia tipicamente durante l'anafase tardiva. Un anello contrattile composto da filamenti di actina si forma appena all'interno della membrana plasmatica, nella precedente piastra metafasica. I filamenti di actina tirano l'equatore della cellula verso l'interno, formando una fessura. Questa fessura è chiamata solco di scissione . Il solco si approfondisce man mano che l'anello di actina si contrae e alla fine la membrana viene scissa in due ( Figura sotto).
Nelle cellule vegetali, una nuova parete cellulare deve formarsi tra le cellule figlie. Durante l'interfase, l'apparato di Golgi accumula enzimi, proteine strutturali e molecole di glucosio prima di rompersi in vescicole e disperdersi nella cellula in divisione. Durante la telofase, queste vescicole di Golgi vengono trasportate su microtubuli per formare un fragmoplasto (una struttura vescicolare) nella piastra metafasica. Lì, le vescicole si fondono e si fondono dal centro verso le pareti cellulari; questa struttura è chiamata piastra cellulare . Man mano che più vescicole si fondono, la piastra cellulare si allarga fino a fondersi con le pareti cellulari alla periferia della cellula. Gli enzimi usano il glucosio che si è accumulato tra gli strati della membrana per costruire una nuova parete cellulare. Le membrane di Golgi diventano parti della membrana plasmatica su entrambi i lati della nuova parete cellulare ( Figura sotto).

La fase G0
modificaNon tutte le cellule aderiscono al classico schema del ciclo cellulare in cui una cellula figlia appena formata entra immediatamente nelle fasi preparatorie dell'interfase, seguite da vicino dalla fase mitotica e dalla citochinesi. Le cellule in fase G 0 non si stanno preparando attivamente a dividersi. La cellula è in una fase quiescente (inattiva) che si verifica quando le cellule escono dal ciclo cellulare. Alcune cellule entrano temporaneamente in G 0 a causa di condizioni ambientali come la disponibilità di nutrienti o la stimolazione da parte di fattori di crescita. La cellula rimarrà in questa fase fino a quando le condizioni non miglioreranno o fino a quando un segnale esterno non innescherà l'inizio di G 1 . Altre cellule che non si dividono mai o raramente, come le cellule muscolari cardiache e nervose mature, rimangono in G0 in modo permanente.
-

-
 Schema che mostra le molecole implicate nella coesione dei cromosomi fratelli
Schema che mostra le molecole implicate nella coesione dei cromosomi fratelli -

-
 Schema del ciclo cellulare animale
Schema del ciclo cellulare animale -

-
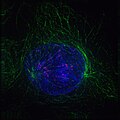 Profase: i microtuboli polari (in verde) formano una struttura intorno al nucleo. I cromosomi, in fase di condensazione, sono in blu, i centromeri in rosso
Profase: i microtuboli polari (in verde) formano una struttura intorno al nucleo. I cromosomi, in fase di condensazione, sono in blu, i centromeri in rosso -
 Prometafase
Prometafase -
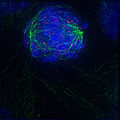 prometafase
prometafase -

-
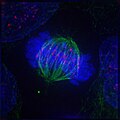 metafase
metafase -

-
 Anafase
Anafase -
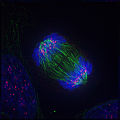 Anafase
Anafase -
 Anafase in cellula di cipolla
Anafase in cellula di cipolla -

-
 Mitosi nelle cellule di cipolla. Profase (in alto) e anafase (in basso)
Mitosi nelle cellule di cipolla. Profase (in alto) e anafase (in basso) -

-

-

-

-




















Il controllo del ciclo cellulare
modificaLa lunghezza del ciclo cellulare è altamente variabile, anche all'interno delle cellule di un singolo organismo. Negli esseri umani, la frequenza del turnover cellulare varia da poche ore nello sviluppo embrionale precoce, a una media di due-cinque giorni per le cellule epiteliali, e a un'intera vita umana trascorsa in G 0 da cellule specializzate, come i neuroni corticali o le cellule del muscolo cardiaco.
C'è anche una variazione nel tempo che una cellula trascorre in ogni fase del ciclo cellulare. Quando le cellule di mammifero in rapida divisione vengono coltivate in una coltura (fuori dal corpo in condizioni di crescita ottimali), la durata del ciclo cellulare è di circa 24 ore. Nelle cellule umane in rapida divisione con un ciclo cellulare di 24 ore, la fase G 1 dura circa nove ore, la fase S dura 10 ore, la fase G 2 dura circa quattro ore e mezza e la fase M dura circa mezz'ora. Per fare un confronto, nelle uova fecondate (e negli embrioni precoci) dei moscerini della frutta, il ciclo cellulare si completa in circa otto minuti. Questo perché il nucleo dell'uovo fecondato si divide molte volte per mitosi ma non passa attraverso la citochinesi finché non è stato prodotto uno "zigote" multinucleato, con molti nuclei situati lungo la periferia della membrana cellulare, accorciando così il tempo del ciclo di divisione cellulare. La tempistica degli eventi nel ciclo cellulare sia degli “invertebrati” che dei “vertebrati” è controllata da meccanismi sia interni che esterni alla cellula.
Regolazione del ciclo cellulare da parte di eventi esterni
modificaSia l'inizio che l'inibizione della divisione cellulare sono innescati da eventi esterni alla cellula quando sta per iniziare il processo di replicazione. Un evento può essere semplice come la morte delle cellule vicine o travolgente come il rilascio di ormoni che promuovono la crescita, come l' ormone della crescita umano (HGH o hGH) . Una mancanza di HGH può inibire la divisione cellulare, con conseguente nanismo, mentre troppo HGH può causare gigantismo. Anche l'affollamento delle cellule può inibire la divisione cellulare. Al contrario, un fattore che può avviare la divisione cellulare è la dimensione della cellula: man mano che una cellula cresce, diventa fisiologicamente inefficiente a causa del suo rapporto superficie-volume in diminuzione. La soluzione a questo problema è dividersi.
Qualunque sia la fonte del messaggio, la cellula riceve il segnale e una serie di eventi al suo interno le consente di procedere in interfase. Andando avanti da questo punto di inizio, ogni parametro richiesto durante ogni fase del ciclo cellulare deve essere soddisfatto o il ciclo non può procedere.
Regolamentazione dei posti di blocco interni
modificaÈ essenziale che le cellule figlie prodotte siano duplicati esatti della cellula madre. Errori nella duplicazione o distribuzione dei cromosomi portano a mutazioni che possono essere trasmesse a ogni nuova cellula prodotta da una cellula anomala. Per impedire che una cellula compromessa continui a dividersi, ci sono meccanismi di controllo interni che operano in tre principali checkpoint del ciclo cellulare : un checkpoint è uno dei diversi punti nel ciclo cellulare eucariotico in cui la progressione di una cellula verso la fase successiva del ciclo può essere interrotta fino a quando le condizioni non sono favorevoli. Questi checkpoint si verificano verso la fine di G 1 , alla transizione G 2 / M e durante la metafase ( Figura sotto).

Il posto di blocco G 1
modificaIl checkpoint G 1 determina se tutte le condizioni sono favorevoli affinché la divisione cellulare proceda. Il checkpoint G 1 , chiamato anche punto di restrizione (nel lievito), è un punto in cui la cellula si impegna irreversibilmente nel processo di divisione cellulare. Influenze esterne, come i fattori di crescita, svolgono un ruolo importante nel portare la cellula oltre il checkpoint G 1. Oltre alle riserve adeguate e alle dimensioni della cellula, c'è un controllo per i danni al DNA genomico al checkpoint G 1. Una cellula che non soddisfa tutti i requisiti non potrà progredire nella fase S. La cellula può interrompere il ciclo e tentare di porre rimedio alla condizione problematica, oppure può avanzare in G 0 e attendere ulteriori segnali quando le condizioni migliorano.
Il posto di blocco G2
modificaIl checkpoint G 2 impedisce l'ingresso nella fase mitotica se non vengono soddisfatte determinate condizioni. Come al checkpoint G 1 , vengono valutate le dimensioni delle cellule e le riserve proteiche. Tuttavia, il ruolo più importante del checkpoint G 2 è quello di garantire che tutti i cromosomi siano stati replicati e che il DNA replicato non sia danneggiato. Se i meccanismi del checkpoint rilevano problemi con il DNA, il ciclo cellulare viene interrotto e la cellula tenta di completare la replicazione del DNA o di riparare il DNA danneggiato.
Il posto di blocco M
modificaIl checkpoint M si verifica verso la fine della fase di metafase della cariocinesi. Il checkpoint M è anche noto come checkpoint del fuso, perché determina se tutti i cromatidi fratelli sono correttamente attaccati ai microtubuli del fuso. Poiché la separazione dei cromatidi fratelli durante l'anafase è una fase irreversibile, il ciclo non procederà finché i cinetocori di ciascuna coppia di cromatidi fratelli non saranno saldamente ancorati ad almeno due fibre del fuso che derivano dai poli opposti della cellula.
Collegamento all'apprendimento
modificaGuarda cosa succede ai checkpoint G1 , G2 e M visitando questo sito web per vedere un'animazione del ciclo cellulare.
Molecole regolatrici del ciclo cellulare
modificaOltre ai checkpoint controllati internamente, ci sono due gruppi di molecole intracellulari che regolano il ciclo cellulare. Queste molecole regolatrici promuovono il progresso della cellula verso la fase successiva (regolazione positiva) o arrestano il ciclo (regolazione negativa). Le molecole regolatrici possono agire individualmente o possono influenzare l'attività o la produzione di altre proteine regolatrici. Pertanto, il fallimento di un singolo regolatore può non avere quasi alcun effetto sul ciclo cellulare, soprattutto se più di un meccanismo controlla lo stesso evento. Tuttavia, l'effetto di un regolatore carente o non funzionante può essere di vasta portata e potenzialmente fatale per la cellula se vengono interessati più processi.
Regolazione positiva del ciclo cellulare
modificaDue gruppi di proteine, chiamate cicline e chinasi ciclina-dipendenti (Cdk), sono definite regolatori positivi. Sono responsabili del progresso della cellula attraverso i vari checkpoint. I livelli delle quattro proteine cicline fluttuano durante il ciclo cellulare secondo uno schema prevedibile ( Figura sotto). Gli aumenti nella concentrazione delle proteine cicline sono innescati da segnali sia esterni che interni. Dopo che la cellula passa alla fase successiva del ciclo cellulare, le cicline che erano attive nella fase precedente vengono degradate dagli enzimi citoplasmatici, come mostrato nella Figura sotto.

Le cicline regolano il ciclo cellulare solo quando sono strettamente legate alle Cdk. Per essere completamente attive, il complesso Cdk/ciclina deve anche essere fosforilato in punti specifici per attivare il complesso. Come tutte le chinasi, le Cdk sono enzimi ( chinasi ) che a loro volta fosforilano altre proteine. La fosforilazione attiva la proteina modificandone la forma. Le proteine fosforilate dalle Cdk sono coinvolte nell'avanzamento della cellula alla fase successiva. ( Figura 10.13 ). I livelli delle proteine Cdk sono relativamente stabili durante tutto il ciclo cellulare; tuttavia, le concentrazioni di ciclina fluttuano e determinano quando si formano i complessi Cdk/ciclina. Le diverse cicline e Cdk si legano in punti specifici del ciclo cellulare e quindi regolano diversi checkpoint.

Poiché le fluttuazioni cicliche dei livelli di ciclina sono in gran parte basate sulla tempistica del ciclo cellulare e non su eventi specifici, la regolazione del ciclo cellulare avviene solitamente tramite le molecole Cdk da sole o tramite i complessi Cdk/ciclina. Senza una concentrazione specifica di complessi ciclina/Cdk completamente attivati, il ciclo cellulare non può procedere attraverso i checkpoint.
Sebbene le cicline siano le principali molecole regolatrici che determinano lo slancio in avanti del ciclo cellulare, ci sono diversi altri meccanismi che regolano con precisione il progresso del ciclo con effetti negativi, piuttosto che positivi. Questi meccanismi bloccano essenzialmente la progressione del ciclo cellulare fino a quando non vengono risolte le condizioni problematiche. Le molecole che impediscono la piena attivazione delle Cdk sono chiamate inibitori delle Cdk. Molte di queste molecole inibitrici monitorano direttamente o indirettamente un particolare evento del ciclo cellulare. Il blocco posto sulle Cdk dalle molecole inibitrici non verrà rimosso fino al completamento dell'evento specifico che l'inibitore monitora.
Regolazione negativa del ciclo cellulare
modificaIl secondo gruppo di molecole regolatrici del ciclo cellulare sono i regolatori negativi , che fermano il ciclo cellulare. Ricorda che nella regolazione positiva, le molecole attive fanno progredire il ciclo.
Le molecole regolatrici negative meglio comprese sono la proteina del retinoblastoma (Rb) , p53 e p21 . Le proteine del retinoblastoma sono un gruppo di proteine oncosoppressori comuni in molte cellule. Dovremmo notare qui che le designazioni 53 e 21 si riferiscono alle masse molecolari funzionali delle proteine (p) in kilodalton (un dalton è uguale a un'unità di massa atomica , che è uguale a un protone o un neutrone o 1 g/mol). Gran parte di ciò che si sa sulla regolazione del ciclo cellulare deriva da ricerche condotte su cellule che hanno perso il controllo regolatorio . Si è scoperto che tutte e tre queste proteine regolatrici erano danneggiate o non funzionali in cellule che avevano iniziato a replicarsi in modo incontrollato (cioè, erano diventate cancerose). In ogni caso, la causa principale del progresso incontrollato attraverso il ciclo cellulare era una copia difettosa della proteina regolatrice.
Rb, p53 e p21 agiscono principalmente al checkpoint G 1. p53 è una proteina multifunzionale che ha un impatto importante sull'impegno di una cellula alla divisione perché agisce quando c'è DNA danneggiato nelle cellule che stanno subendo i processi preparatori durante G 1. Se viene rilevato DNA danneggiato, p53 interrompe il ciclo cellulare e quindi recluta enzimi specifici per riparare il DNA. Se il DNA non può essere riparato, p53 può innescare l'apoptosi, o suicidio cellulare, per prevenire la duplicazione dei cromosomi danneggiati. Quando i livelli di p53 aumentano, viene innescata la produzione di p21. p21 impone l'arresto del ciclo dettato da p53 legandosi e inibendo l'attività dei complessi Cdk/ciclina. Quando una cellula è esposta a più stress, si accumulano livelli più elevati di p53 e p21, rendendo meno probabile che la cellula passi alla fase S.
Rb, che controlla ampiamente le dimensioni delle cellule, esercita la sua influenza regolatrice su altre proteine regolatrici positive. Nello stato attivo , defosforilato, Rb si lega a proteine chiamate fattori di trascrizione , più comunemente E2F ( Figura 10.14 ). I fattori di trascrizione "accendono" geni specifici, consentendo la produzione di proteine codificate da quel gene. Quando Rb è legato a E2F, la produzione di proteine necessarie per la transizione G 1 /S viene bloccata. Man mano che la cellula aumenta di dimensioni, Rb viene lentamente fosforilata fino a diventare inattivata . Rb rilascia E2F, che ora può accendere il gene che produce la proteina di transizione, e questo particolare blocco viene rimosso. Affinché la cellula superi ciascuno dei checkpoint, tutti i regolatori positivi devono essere "accesi" e tutti i regolatori negativi devono essere "spenti".

Cancro e ciclo cellulare
modificaIl cancro comprende molte malattie diverse causate da un meccanismo comune: la crescita incontrollata delle cellule. Nonostante la ridondanza e i livelli sovrapposti di controllo del ciclo cellulare, si verificano degli errori. Uno dei processi critici monitorati dal meccanismo di sorveglianza del checkpoint del ciclo cellulare è la corretta replicazione del DNA durante la fase S. Anche quando tutti i controlli del ciclo cellulare sono completamente funzionali, una piccola percentuale di errori di replicazione (mutazioni) verrà trasmessa alle cellule figlie. Se si verificano modifiche alla sequenza nucleotidica del DNA all'interno di una porzione codificante di un gene e non vengono corrette, si verifica una mutazione genetica. Tutti i tumori iniziano quando una mutazione genetica dà origine a una proteina difettosa che svolge un ruolo chiave nella riproduzione cellulare.
Il cambiamento nella cellula che deriva dalla proteina malformata può essere minore: forse un leggero ritardo nel legame di Cdk alla ciclina o una proteina Rb che si stacca dal suo DNA bersaglio mentre è ancora fosforilata. Tuttavia, anche errori minori possono consentire che errori successivi si verifichino più facilmente. Più e più volte, piccoli errori non corretti vengono trasmessi dalla cellula madre alle cellule figlie e amplificati poiché ogni generazione produce più proteine non funzionali da danni al DNA non corretti. Alla fine, il ritmo del ciclo cellulare accelera man mano che l'efficacia dei meccanismi di controllo e riparazione diminuisce. La crescita incontrollata delle cellule mutate supera la crescita delle cellule normali nell'area e può verificarsi un tumore ("-oma").

Proto-oncogeni
modificaI geni che codificano per i regolatori positivi del ciclo cellulare sono chiamati proto-oncogeni . I proto-oncogeni sono geni normali che, quando mutati in certi modi, diventano oncogeni , geni che causano la cancerizzazione di una cellula. Considera cosa potrebbe accadere al ciclo cellulare in una cellula con un oncogene acquisito di recente. Nella maggior parte dei casi, l'alterazione della sequenza del DNA si tradurrà in una proteina meno funzionale (o non funzionale). Il risultato è dannoso per la cellula e probabilmente impedirà alla cellula di completare il ciclo cellulare; tuttavia, l'organismo non viene danneggiato perché la mutazione non verrà portata avanti. Se una cellula non può riprodursi, la mutazione non viene propagata e il danno è minimo. Occasionalmente, tuttavia, una mutazione genetica provoca un cambiamento che aumenta l'attività di un regolatore positivo. Ad esempio, una mutazione che consente a Cdk di essere attivato senza essere associato alla ciclina potrebbe spingere il ciclo cellulare oltre un checkpoint prima che tutte le condizioni richieste siano soddisfatte. Se le cellule figlie risultanti sono troppo danneggiate per subire ulteriori divisioni cellulari, la mutazione non si propagherebbe e non si verificherebbe alcun danno all'organismo. Tuttavia, se le cellule figlie atipiche sono in grado di subire ulteriori divisioni cellulari, le generazioni successive di cellule potrebbero accumulare ancora più mutazioni, alcune possibilmente in geni aggiuntivi che regolano il ciclo cellulare.
Il gene Cdk nell'esempio sopra è solo uno dei tanti geni che sono considerati proto-oncogeni. Oltre alle proteine regolatrici del ciclo cellulare, qualsiasi proteina che influenza il ciclo può essere alterata in modo tale da ignorare i checkpoint del ciclo cellulare. Un oncogene è qualsiasi gene che, quando alterato, porta a un aumento del tasso di progressione del ciclo cellulare.
Geni soppressori del tumore
modificaCome i proto-oncogeni, molte delle proteine regolatrici negative del ciclo cellulare sono state scoperte in cellule che erano diventate cancerose. I geni oncosoppressori sono segmenti di DNA che codificano per proteine regolatrici negative, il tipo di regolatori che, quando attivati, possono impedire alla cellula di subire una divisione incontrollata. La funzione collettiva delle proteine del gene oncosoppressore meglio comprese, Rb, p53 e p21, è quella di creare un ostacolo alla progressione del ciclo cellulare finché non si siano completati determinati eventi. Una cellula che trasporta una forma mutata di un regolatore negativo potrebbe non essere in grado di fermare il ciclo cellulare se c'è un problema. I soppressori oncologici sono simili ai freni di un veicolo: freni malfunzionanti possono contribuire a un incidente d'auto!
I geni p53 mutati sono stati identificati in oltre il 50 percento di tutte le cellule tumorali umane. Questa scoperta non sorprende alla luce dei molteplici ruoli che la proteina p53 svolge al checkpoint G 1. Una cellula con un p53 difettoso potrebbe non riuscire a rilevare gli errori presenti nel DNA genomico ( Figura 10.16 ). Anche se un p53 parzialmente funzionale identifica le mutazioni, potrebbe non essere più in grado di segnalare gli enzimi di riparazione del DNA necessari. In entrambi i casi, il DNA danneggiato rimarrà non corretto. A questo punto, un p53 funzionale considererà la cellula non recuperabile e innescherà la morte cellulare programmata (apoptosi). La versione danneggiata di p53 trovata nelle cellule tumorali, tuttavia, non può innescare l'apoptosi.

| Esercizio |
|---|
Il papillomavirus umano può causare il cancro cervicale. Il virus codifica E6, una proteina che lega p53. Sulla base di questo fatto e di ciò che sai su p53, quale effetto pensi che abbia il legame E6 sull'attività di p53?
|
La perdita della funzione p53 ha altre ripercussioni sul ciclo cellulare. Il p53 mutato potrebbe perdere la sua capacità di innescare la produzione di p21. Senza livelli adeguati di p21, non c'è un blocco efficace sull'attivazione di Cdk. In sostanza, senza un p53 completamente funzionale, il checkpoint G 1 è gravemente compromesso e la cellula procede direttamente da G 1 a S indipendentemente dalle condizioni interne ed esterne. Al completamento di questo ciclo cellulare abbreviato, vengono prodotte due cellule figlie che hanno ereditato il gene p53 mutato. Date le condizioni non ottimali in cui la cellula madre si è riprodotta, è probabile che le cellule figlie abbiano acquisito altre mutazioni oltre al gene oncosoppressore difettoso. Cellule come queste cellule figlie accumulano rapidamente sia oncogeni che geni oncosoppressori non funzionali. Ancora una volta, il risultato è la crescita del tumore.
La divisione cellulare procariotica
modificaI procarioti, come i batteri, producono cellule figlie tramite fissione binaria. Per gli organismi unicellulari, la divisione cellulare è l'unico metodo per produrre nuovi individui. Sia nelle cellule procariotiche che in quelle eucariotiche, il risultato della riproduzione cellulare è una coppia di cellule figlie che sono geneticamente identiche alla cellula madre. Negli organismi unicellulari, le cellule figlie sono individui.
Per ottenere il risultato di una prole clonata, sono essenziali alcuni passaggi. Il DNA genomico deve essere replicato e quindi allocato nelle cellule figlie; anche il contenuto citoplasmatico deve essere diviso per dare a entrambe le nuove cellule il macchinario cellulare per sostenere la vita. Come abbiamo visto con le cellule batteriche, il genoma è costituito da un singolo cromosoma di DNA circolare; pertanto, il processo di divisione cellulare è semplificato. La cariocinesi è inutile perché non c'è un vero nucleo e quindi non c'è bisogno di dirigere una copia dei cromosomi multipli in ogni cellula figlia. Questo tipo di divisione cellulare è chiamata fissione binaria (procariotica) .
Scissione binaria
modificaGrazie alla relativa semplicità dei procarioti, il processo di divisione cellulare è un processo meno complicato e molto più rapido della divisione cellulare negli eucarioti. Come ripasso delle informazioni generali sulla divisione cellulare di cui abbiamo parlato all'inizio di questo capitolo, ricorda che il singolo cromosoma circolare del DNA dei batteri occupa una posizione specifica, la regione del nucleoide, all'interno della cellula ( Figura sotto). Sebbene il DNA del nucleoide sia associato a proteine che aiutano a impacchettare la molecola in una dimensione compatta, non ci sono proteine istoniche e quindi nessun nucleosoma nei procarioti. Le proteine di impacchettamento dei batteri sono, tuttavia, correlate alle proteine coesina e condensina coinvolte nella compattazione cromosomica degli eucarioti.


Il cromosoma batterico è attaccato alla membrana plasmatica circa a metà della cellula. Il punto di partenza della replicazione, l' origine , è vicino al sito di legame del cromosoma alla membrana plasmatica ( Figura sopra). La replicazione del DNA è bidirezionale, allontanandosi dall'origine su entrambi i filamenti dell'ansa simultaneamente. Quando si formano i nuovi doppi filamenti, ogni punto di origine si allontana dall'attacco della parete cellulare verso le estremità opposte della cellula. Quando la cellula si allunga, la membrana in crescita aiuta il trasporto dei cromosomi. Dopo che i cromosomi hanno superato il punto medio della cellula allungata, inizia la separazione citoplasmatica. La formazione di un anello composto da unità ripetute di una proteina chiamata FtsZ (abbreviazione di "filamenting temperature-sensitive mutant Z") dirige la partizione tra i nucleoidi. La formazione dell'anello FtsZ innesca l'accumulo di altre proteine che lavorano insieme per reclutare nuovi materiali della membrana e della parete cellulare nel sito. Si forma un setto tra i nucleoidi figli, che si estende gradualmente dalla periferia verso il centro della cellula. Quando le nuove pareti cellulari sono in posizione, le cellule figlie si separano.